Как именно энергия запасается в АТФ (аденозинтрифосфат), и как она отдается для совершения какой-то полезной работы? Кажется невероятно сложным, что некая абстрактная энергия вдруг получает материальный носитель в виде молекулы, находящейся внутри живых клеток, и что она может высвобождаться не в виде тепла (что более-менее понятно), а в виде создания другой молекулы. Обычно авторы учебников ограничиваются фразой «энергия запасается в виде высокоэнергетической связи между частями молекулы, и отдается при разрыве этой связи, совершая полезную работу», но это ничего не объясняет.

В самых общих чертах эти манипуляции с молекулами и энергией происходят так: сначала . Или создаются в хлоропластах в цепи похожих реакций. На это тратится энергия, получаемая при контролируемом сгорании питательных веществ прямо внутри митохондрий или энергия фотонов солнечного света, падающих на молекулу хлорофилла. Потом АТФ доставляется в те места клетки, где необходимо совершить какую-то работу. И при отщеплении от нее одной или двух фосфатных групп выделяется энергия, которая эту работу и совершает. АТФ при этом распадается на две молекулы: если отщепилась только одна фосфатная группа, то АТФ превращается в АДФ (аденозинДИфосфат, отличающийся от аденозинТРИфосфата только отсутствием той самой отделившейся фосфатной группы). Если АТФ отдала сразу две фосфатные группы, то энергии выделяется больше, а от АТФ остается аденозинМОНОфосфат (АМФ ).
Очевидно, что клетке необходимо осуществлять и обратный процесс, превращая молекулы АДФ или АМФ в АТФ, чтобы цикл мог повториться. Но эти молекулы-«заготовки» могут спокойно плавать рядом с недостающими им для превращения в АТФ фосфатами, и никогда с ними не объединиться, потому что такая реакция объединения энергетически невыгодна.

Что такое «энергетическая выгода» химической реакции, понять довольно просто, если знать о втором законе термодинамики : во Вселенной или в любой системе, изолированной от остальных, беспорядок может лишь нарастать. То есть сложноорганизованные молекулы, сидящие в клетке в чинном порядке, в соответствии с этим законом могут только разрушаться, образуя более мелкие молекулы или даже распадаясь на отдельные атомы, ведь тогда порядка будет заметно меньше. Чтобы понять эту мысль, можно сравнить сложную молекулу с собранным из Лего самолетиком. Тогда мелкие молекулы, на которые распадается сложная, будут ассоциироваться с отдельными частями этого самолета, а атомы — с отдельными кубиками Лего. Посмотрев на аккуратно собранный самолет и сравнив его с беспорядочной кучей деталей, становится понятно, почему сложные молекулы содержат больше порядка, чем мелкие.
Такая реакция распада (молекул, не самолета) будет энергетически выгодной, а значит может осуществляться самопроизвольно, и при распаде будет выделяться энергия. Хотя на самом деле и расщепление самолета будет энергетически выгодно: несмотря на то, что сами по себе детали отщепляться друг от друга не будут и над их отцеплением придется попыхтеть сторонней силе в виде пацана, который хочет использовать эти детали для чего-то другого, он затратит на превращение самолета в хаотическую кучу деталей энергию, полученную от поедания высокоупорядоченной пищи. И чем плотнее слиплись детали, тем больше энергии будет потрачено, в том числе выделено в виде тепла. Итог: кусок плюшки (источник энергии) и самолет превращены в беспорядочную массу, молекулы воздуха вокруг ребенка нагрелись (а значит движутся более беспорядочно) — хаоса стало больше, то есть расщепление самолета энергетически выгодно.
Подводя итог, можно сформулировать такие правила, следующие из второго закона термодинамики:
1. При снижении количества порядка энергия выделяется, происходят энергетически выгодные реакции
2. При увеличении количества порядка энергия поглощается, происходят энергетически затратные реакции
На первый взгляд, такое неизбежное движение от порядка к хаосу делает невозможным обратные процессы, такие как построение из одной оплодотворенной яйцеклетки и молекул питательных веществ, поглощеных матерью-коровой, несомненно весьма упорядоченного по сравнению с пережеванной травой теленка.
Но все-таки это происходит, и причина этого в том, что живые организмы имеют одну фишку, позволяющую и поддержать стремление Вселенной к энтропии, и построить себя и свое потомство: они объединяют в один процесс две реакции, одна из которых энергетически выгодна, а другая энергозатратна . Таким совмещением двух реакций можно добиться того, чтобы энергия, выделяемая при первой реакции, с избытком перекрывала энергетические затраты второй. В примере с самолетом отдельно взятое его разбирание энергозатратно, и без стороннего источника энергии в виде разрушенной метаболизмом пацана плюшки самолет стоял бы вечно.

Это как при катании с горки на санках: сначала человек во время поглощения пищи запасает энергию, полученную в результате энергетически выгодных процессов расщепления высокоупорядоченной курицы на молекулы и атомы в его организме. А потом тратит эту энергию, затаскивая санки на гору. Перемещение санок от подножия к вершине энергетически невыгодно, поэтому самопроизвольно они туда никогда не закатятся, на это нужна какая-то сторонняя энергия. И если энергии, полученной от поедания курицы, будет недостаточно для преодоления подъема, то процесса «скатывание на санках с вершины горы» не будет.
Именно энергозатратные реакции (energy-consuming reaction ) увеличивают количество порядка, поглощая энергию, выделяемую при сопряженной реакции. И баланс между выделением и потреблением энергии в этих сопряженных реакциях всегда должен быть положительным, то есть их совокупность будет увеличивать количество хаоса. Примером увеличения энтропии (неупорядоченности) (entropy [‘entrə pɪ ] ) является выделение тепла при энергодающей реакции (energy supply reaction ): соседние с вступившими в реакцию молекулами частицы вещества получают энергичные толчки от реагирующих, начинают двигаться быстрее и хаотичнее, распихивая в свою очередь другие молекулы и атомы этого и соседних веществ.

Вернемся еще раз к получению энергии из пищи: кусок Banoffee Pie гораздо более упорядочен, чем получившаяся в результате пережевывания масса, попавшая в желудок. Которая в свою очередь состоит из крупных, более упорядоченных молекул, чем те, на которые ее расщепит кишечник. А они в свою очередь будут доставлены в клетки тела, где от них будут отрывать уже отдельные атомы и даже электроны… И на каждом этапе увеличения хаоса в отдельно взятом куске торта будет происходить выделение энергии, которую улавливают органы и органеллы счастливого поедателя, запасая ее в виде АТФ (энергозатратно), пуская на построение новых нужных молекул (энергозатратно) или на нагревание тела (тоже энергозатратно). В системе «человек — Banoffee Pie — Вселенная» порядка в результате этого стало меньше (за счет разрушения кейка и выделения тепловой энергии перерабатывающими его органеллами), но в отдельно взятом человеческом теле счастья порядка стало больше (за счет возникновения новых молекул, частей органелл и целых клеточных органов).
Если вернуться к молекуле АТФ, после всего этого термодинамического отступления становится понятно, что на создание ее из составных частей (более мелких молекул) необходимо затратить энергию, полученную от энергетически выгодных реакций. Один из способов ее создания подробно описан , другой (весьма схожий) используется в хлоропластах, где вместо энергии протонного градиента используется энергия фотонов, испущенных Солнцем.

Можно выделить три группы реакций, в результате которых производится АТФ (смотри схему справа):
- расщепление глюкозы и жирных кислот на крупные молекулы в цитоплазме уже позволяет получить некоторое количество АТФ (небольшое, на одну расщепленную на этом этапе молекулу глюкозы приходится всего лишь 2 полученные молекулы АТФ). Но основная цель этого этапа заключается в создании молекул, использующихся в дыхательной цепи митохондрий.
- дальнейшее расщепление полученных на предыдущем этапе молекул в цикле Кребса, протекающее в матриксе митохондрий, дает всего одну молекулу АТФ, его основная цель та же, что и в прошлом пункте.
- наконец накопленные на предыдущих этапах молекулы используются в дыхательной цепи митохондрий для производства АТФ, и вот тут его выделяется много (про это подробнее ниже).
Если описать все это более развернуто, взглянув на те же реакции с точки зрения получения и затрат энергии, получится вот что:
0. Молекулы пищи аккуратно сжигаются (окисляются) в первичном расщеплении, происходящем в цитоплазме клетки, а также в цепи химических реакций под названием «цикл Кребса», протекающем уже в матриксе митохондрий — энергодающая часть подготовительного этапа.
В результате сопряжения с этими энергетически выгодными реакциями других, уже энергетически невыгодных реакций создания новых молекул образуются 2 молекулы АТФ и несколько молекул других веществ — энергозатратная часть подготовительного этапа. Эти попутно образующиеся молекулы являются переносчиками высокоэнергетических электронов, которые будут использованы в дыхательной цепи митохондрий на следующем этапе.

1. На мембранах митохондрий, бактерий и некоторых архей происходит энергодающее отщепление протонов и электронов от молекул, полученных в предыдущем этапе (но не от АТФ). Прохождение электронов по комплексам дыхательной цепи (I, III и IV на схеме слева) показано желтыми извилистыми стрелками, прохождение через эти комплексы (а значит, и через внутреннюю мембрану митохондрии) протонов — красными стрелками.
Почему электроны нельзя просто отщепить от молекулы-переносчика с использованием мощного окислителя-кислорода и использовать выделяющуюся энергию? Зачем передавать их от одного комплекса к другому, ведь в итоге они к тому же кислороду и приходят? Оказывается, чем больше разница в способности притягивать электроны у электронодающей (восстановителя ) и электроноберущей (окислителя ) молекул, участвующих в реакции передачи электрона, тем большая энергия выделяется при этой реакции.
Разница в такой способности у образующихся в цикле Кребса молекул-переносчиков электронов и кислорода такова, что выделившейся при этом энергии было бы достаточно для синтеза нескольких молекул АТФ. Но из-за такого резкого перепада в энергии системы эта реакция протекала бы с почти взрывной мощью, и почти вся энергия выделялась бы в виде неулавливаемого тепла, то есть фактически терялась.

Живые клетки же делят эту реакцию на несколько маленьких стадий, сначала передавая электроны от слабо притягивающих молекул-носителей к чуть сильнее притягивающему первому комплексу в дыхательной цепи, от него к еще немного сильнее притягивающему убихинону (или коэнзиму Q-10 ), чья задача заключается в перетаскивании электронов к следующему, еще немного сильнее притягивающему дыхательному комплексу, который получает свою часть энергии от этого несостоявшегося взрыва, пуская ее на прокачку протонов через мембрану.. И так до момента, пока электроны не встретятся наконец с кислородом, притянувшись к нему, прихватив пару протонов, и не образуют молекулу воды. Такое деление одной мощной реакции на мелкие шаги позволяет почти половину полезной энергии направить на совершение полезной работы: в данном случае на создание протонного электрохимического градиента , о котором речь пойдет во втором пункте.
Как именно энергия передаваемых электронов помогает сопряженной энергозатратной реакции прокачки протонов через мембрану, сейчас только начинают выяснять. Скорее всего, присутствие электрически заряженной частицы (электрона) влияет на конфигурацию того места во встроенном в мембрану протеине, где он находится: так, что это изменение провоцирует затягивание протона в протеин и его движение через протеиновый канал в мембране. Важно то, что фактически энергия, полученная в результате отщепления высокоэнергетичных электронов от молекулы-носителя и итоговой передачи их кислороду, запасается в виде протонного градиента.
2. Энергия протонов, накопившихся в результате событий из пункта 1 с внешней стороны мембраны и стремящихся попасть на внутреннюю сторону, состоит из двух однонаправленных сил:
- электрической (положительный заряд протонов стремится перейти в место скопления отрицательных зарядов с другой стороны мембраны) и
- химической (как в случае любых других веществ, протоны пытаются равномерно рассеяться в пространстве, распространившись из мест с их высокой концентрацией в места, где их мало)

Электрическое притяжение протонов к отрицательно заряженной стороне внутренней мембраны является намного более мощной силой, чем возникающее из-за разницы в концентрации протонов их стремление перейти в место с меньшей концентрацией (это обозначено шириной стрелок на схеме вверху). Совместная энергия этих влекущих сил настолько велика, что ее хватает и на перемещение протонов внутрь мембраны, и на подпитывание сопутствующей энергозатратной реакции: создание АТФ из АДФ и фосфата.
Рассмотрим подробнее, почему на это нужна энергия, и как именно энергия стремления протонов превращается в энергию химической связи между двумя частями молекулы АТФ.

Молекула АДФ (на схеме справа) не жаждет обзаводиться еще одной фосфатной группой: тот атом кислорода, к которому эта группа может прикрепиться, заряжен так же отрицательно, как и фосфат, а значит они взаимно отталкиваются. И вообще АДФ не собирается вступать в реакции, она химически пассивна. У фосфата, в свою очередь, к тому атому фосфора, который мог бы стать местом связи фосфата и АДФ при создании молекулы АТФ, присоединен собственный атом кислорода, так что и он инициативы проявить не может.
Поэтому эти молекулы необходимо связать одним ферментом, развернуть их так, чтобы связи между ними и «лишними» атомами ослабли и разорвались, а после этого подвести два химически активных конца этих молекул, на которых атомы испытывают недостаток и избыток электронов, друг к другу.
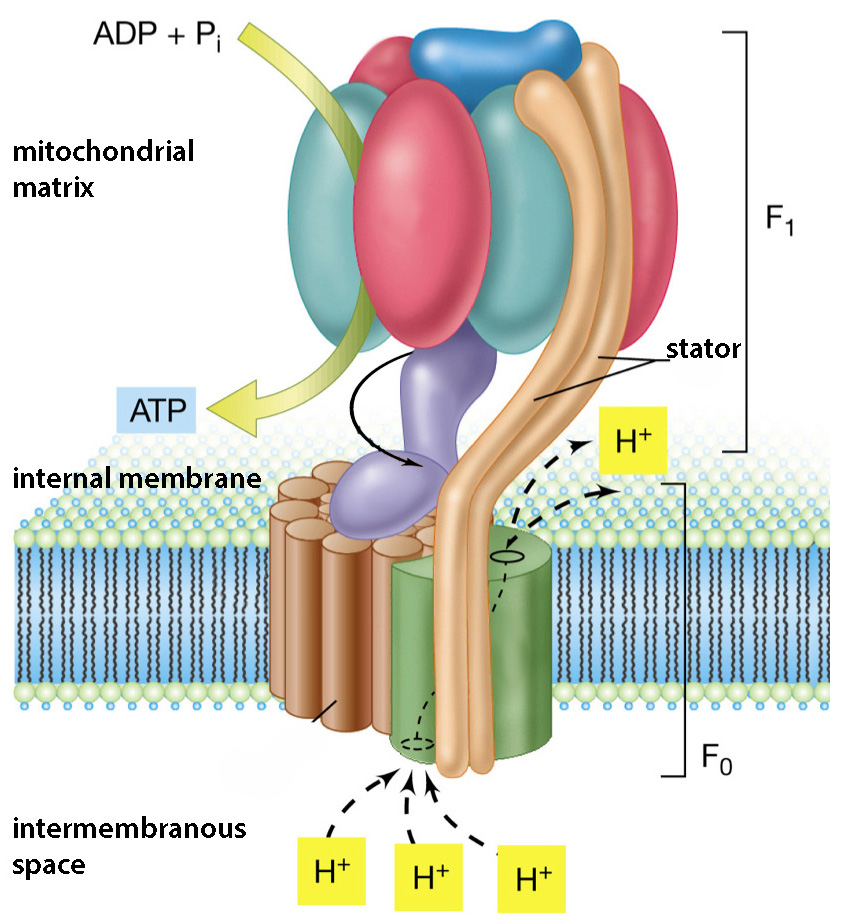
Попавшие в поле взаимной досягаемости ионы фосфора (P +) и кислорода (O —) связываются прочной ковалентной связью за счет того, что совместно овладевают одним электроном, изначально принадлежавшим кислороду. Этим обрабатывающим молекулы ферментом является АТФ-синтаза , а энергию на изменение и своей конфигурации, и взаимного расположения АДФ и фосфата она получает от проходящих через нее протонов. Протонам энергетически выгодно попасть на противоположно заряженную сторону мембраны, где к тому же их мало, а единственный путь проходит через фермент, «ротор» которого протоны попутно вращают.
Строение АТФ-синтазы показано на схеме справа. Ее вращающийся за счет прохождения протонов элемент выделен фиолетовым цветом, а на подвижной картинке внизу показана схема его вращения и создания при этом молекул АТФ. Фермент работает практически как молекулярный мотор, превращая электрохимическую энергию тока протонов в механическую энергию трения двух наборов протеинов друг о друга: вращающаяся «ножка» трется о неподвижные протеины «шляпки гриба», при этом субъединицы «шляпки» изменяют свою форму. Эта механическая деформация превращается в энергию химических связей при синтезе АТФ, когда молекулы АДФ и фосфата обрабатываются и разворачиваются нужным для образования между ними ковалентной связи образом.

Каждая АТФ-синтаза способна синтезировать до 100 молекул АТФ в секунду, и на каждую синтезируемую молекулу АТФ через синтетазу должно пройти около трех протонов. Большая часть синтезируемых в клетках АТФ образуется именно этим путем, и лишь небольшая часть является результатом первичной обработки молекул пищи, происходящей вне митохондрий.
В любой момент в типичной живой клетке находится примерно миллиард молекул АТФ. Во многих клетках вся эта АТФ сменяется (т.е. используется и создается вновь) каждые 1-2 минуты. Средний человек в состоянии покоя использует каждые 24 часа массу АТФ, примерно равную его собственной массе.

В целом почти половина энергии, выделяющаяся при окислении глюкозы или жирных кислот до углекислого газа и воды, улавливается и используется для протекания энергетически невыгодной реакции образования АТФ из АДФ и фосфатов. Коэффициент полезного действия в размере 50% — это очень неплохо, например двигатель автомобиля пускает на полезную работу всего лишь 20% содержащейся в топливе энергии. При этом остальная энергия в обоих случаях рассеивается в виде тепла, и так же как некоторые автомобили, животные постоянно тратят этот избыток (хоть и не полностью, конечно) на разогревание тела. В процессе упомянутых здесь реакций одна молекула глюкозы, постепенно расщепленная до углекислого газа и воды, поставляет клетке 30 молекул АТФ.
Итак, с тем, откуда берется энергия и как именно она запасается в АТФ, все более-менее понятно. Осталось понять, как именно запасенная энергия отдается и что при этом происходит на молекулярно-атомном уровне.

Образованная ковалентная связь между АДФ и фосфатом называется высокоэнергетичной по двум причинам:
- при ее разрушении выделяется много энергии
- электроны, участвующие в создании этой связи (то есть вращающиеся вокруг атомов кислорода и фосфора, между которыми эта связь образована) высокоэнергетичны, то есть находятся на «высоких» орбитах вокруг ядер атомов. И им было бы энергетически выгодно перескочить на уровень пониже, выделив излишек энергии, но пока они находятся именно в этом месте, скрепляя атомы кислорода и фосфора, «спрыгнуть» не получится.
Это стремление электронов упасть на более удобную низкоэнергетичную орбиту обеспечивает и легкость разрушения высокоэнергетичной связи, и выделяемую при этом в виде фотона (являющегося переносчиком электромагнитного взаимодействия) энергию. В зависимости от того, какие молекулы будут подставлены ферментами к разрушающейся молекуле АТФ, какая именно молекула поглотит испущенный электроном фотон, могут происходить разные варианты событий. Но каждый раз энергия, запасенная в виде высокоэнергетической связи, будет использоваться на какие-то нужды клетки :
Сценарий 1: фосфат может быть перенесен на молекулу другого вещества. При этом высокоэнергетичные электроны образуют новую связь, уже между фосфатом и крайним атомом этой молекулы-реципиента. Условием протекания такой реакции является ее энергетическая выгода: в этой новой связи электрон должен обладать немного меньшей энергией, чем когда он был частью молекулы АТФ, испустив часть энергии в виде фотона вовне.

Цель такой реакции заключается в активации молекулы-рецепиента (на схеме слева она обозначена В -ОН): до присоединения фосфата она была пассивной и не могла вступить в реакцию с другой пассивной молекулой А , но теперь она является обладателем запаса энергии в виде высокоэнергетичного электрона, а значит может ее куда-то потратить. Например, на то, чтобы присоединить к себе молекулу А , которую без такого финта ушами (то есть высокой энергии связующего электрона) присоединить невозможно. Фосфат при этом отсоединяется, сделав свое дело.
Получается такая цепочка реакций:
1. АТФ + пассивная молекула В ➡️ АДФ + активная за счет присоединенного фосфата молекула В-Р
2. активированная молекула В-Р + пассивная молекула А ➡️соединенные молекулы А-В + отщепившийся фосфат (Р )
Обе этих реакции энергетически выгодные: в каждой из них участвует высокоэнергетичный связующий электрон, который при разрушении одной связи и построении другой теряет часть своей энергии в виде испускания фотона. В результате этих реакций соединились две пассивные молекулы. Если рассмотреть реакцию соединения этих молекул напрямую (пассивная молекула В + пассивная молекула А ➡️соединенные молекулы А-В ), то она оказывается энергетически затратной, и совершиться не может. Клетки «совершают невозможное», сопрягая эту реакцию с энергетически выгодной реакцией расщепления АТФ на АДФ и фосфат во время совершения тех двух реакций, которые описаны выше. Отщепление происходит в два этапа, на каждом из которых часть энергии связующего электрона тратится на совершение полезной работы, а именно на создание нужных связей между двумя молекулами, из которых получается третья (А-В ), необходимая для функционирования клетки.
Сценарий 2: фосфат может быть отщеплен одномоментно от молекулы АТФ, а выделяющаяся энергия улавливается ферментом или рабочим протеином и тратится на совершение полезной работы.

Как можно уловить что-то настолько неощутимое, как ничтожное возмущение электромагнитного поля в момент падения электрона на более низкую орбиту? Очень просто: с помощью других электронов и с помощью атомов, способных поглотить выделяемый при этом электроном фотон.

Атомы, составляющие молекулы, скреплены в прочные цепочки и кольца за счет (такую цепочку представляет собой несвернутый протеин на картинке справа). А отдельные части этих молекул притянуты друг к другу более слабыми электромагнитными взаимодействиями (например, водородными связями или силами Ван дер Ваальса), что и позволяет им сфорачиваться в сложные структуры. Некоторые из этих конфигураций атомов очень стабильны, и никакое возмущение электромагнитного поля их не поколебит.. не поколебёт.. в общем, они устойчивы. А некоторые довольно подвижны, и достаточно легкого электромагнитного пинка, чтобы они изменили свою конфигурацию (обычно это не ковалентные связи). И именно такой пинок дает им тот самый прилетевший фотон-переносчик электромагнитного поля, испущенный перешедшим на более низкую орбиту электроном при отсоединении фосфата.
Изменения конфигурации протеинов в результате расщепления молекул АТФ ответственны за самые удивительные события, происходящие в клетке. Наверняка те, кто интересуются клеточными процессами хотя бы на уровне «посмотрю их анимацию на youtube» натыкались на видео, показывающее протеиновую молекулу кинезина , в прямом смысле слова шагающую, переставляя ноги, по нити клеточного скелета, перетаскивая присоединенный к ней груз.
Именно отщепление фосфата от АТФ обеспечивает это шагание, и вот каким образом:

Кинезин (kinesin ) относится к особому виду протеинов, которым свойственно спонтанно менять свою конформацию (взаимное положение атомов в молекуле). Оставленный в покое, он случайным образом переходит из конформации 1, в которой он прикреплен одной «ногой» к актиновому филаменту (actin filament ) — самой тонкой нити, образующей цитоскелет клетки (cytoskeleton ), в конформацию 2, сделав таким образом шаг вперед и стоя на двух «ногах». Из конформации 2 он с равной вероятностью перейдет как в конформацию 3 (приставляет заднюю ногу к передней), так и обратно в конформацию 1. Поэтому движения кинезина в каком-либо направлении не происходит, он просто бесцельно фланирует.

Но все меняется, стоит ему соединиться с молекулой АТФ. Как показано на схеме слева, присоединение АТФ к кинезину, находящемуся в конформации 1, приводит к изменению его пространственного положения и он переходит в конформацию 2. Причина этого — взаимное электромагнитное влияние молекул АТФ и кинезина друг на друга. Эта реакция является обратимой, потому что энергии затрачено не было, и если АТФ отсоединится от кинезина, он просто поднимет «ногу», оставшись на месте, и будет ждать следующую молекулу АТФ.
Но если она задержится, то из-за взаимного притяжения этих молекул связь, удерживающая фосфат в пределах АТФ, разрушается. Выделившаяся при этом энергия, а так же распад АТФ на две молекулы (которые уже по другому влияют своими электромагнитными полями на атомы кинезина) приводят к тому, что конформация кинезина меняется: он «подтаскивает заднюю ногу». Осталось сделать шаг вперед, что и происходит при отсоединении АДФ и фосфата, возвращающем кинезин в исходную конформацию 1.
В результате гидролиза АТФ кинезин сдвинулся вправо, и как только к нему присоединится следующая молекула, он сделает еще одну пару шагов, использовав запасенную в ней энергию.
Важно, что кинезин, находящийся в конформации 3 с присоединенными АДФ и фосфатом не может вернуться в конформацию 2, сделав «шаг назад». Это объясняется все тем же принципом соответствия второму закону терморегуляции: переход системы «кинезин + АТФ» из конформации 2 в конформацию 3 сопровождается выделением энергии, а значит обратный переход будет энергозатратным. Чтобы он произошел, нужно откуда-то взять энергию на соединение АДФ с фосфатом, а взять ее в этой ситуации неоткуда. Поэтому соединенному с АТФ кинезину открыт путь только в одну сторону, что и позволяет совершать полезную работу по перетаскиванию чего-либо из одного конца клетки в другой. Кинезин например участвует в растаскивании хромосом делящейся клетки при митозе (процессе деления эукариотических клеток). А мышечный протеин миозин бежит вдоль актиновых филаментов, вызывая сокращение мышцы.

Это движение бывает очень быстрым: некоторые моторные (отвечающие за различные формы клеточной подвижности) протеины, задействованные в репликации генов, мчатся вдоль цепочки ДНК со скоростью тысячи нуклеотидов в секунду.
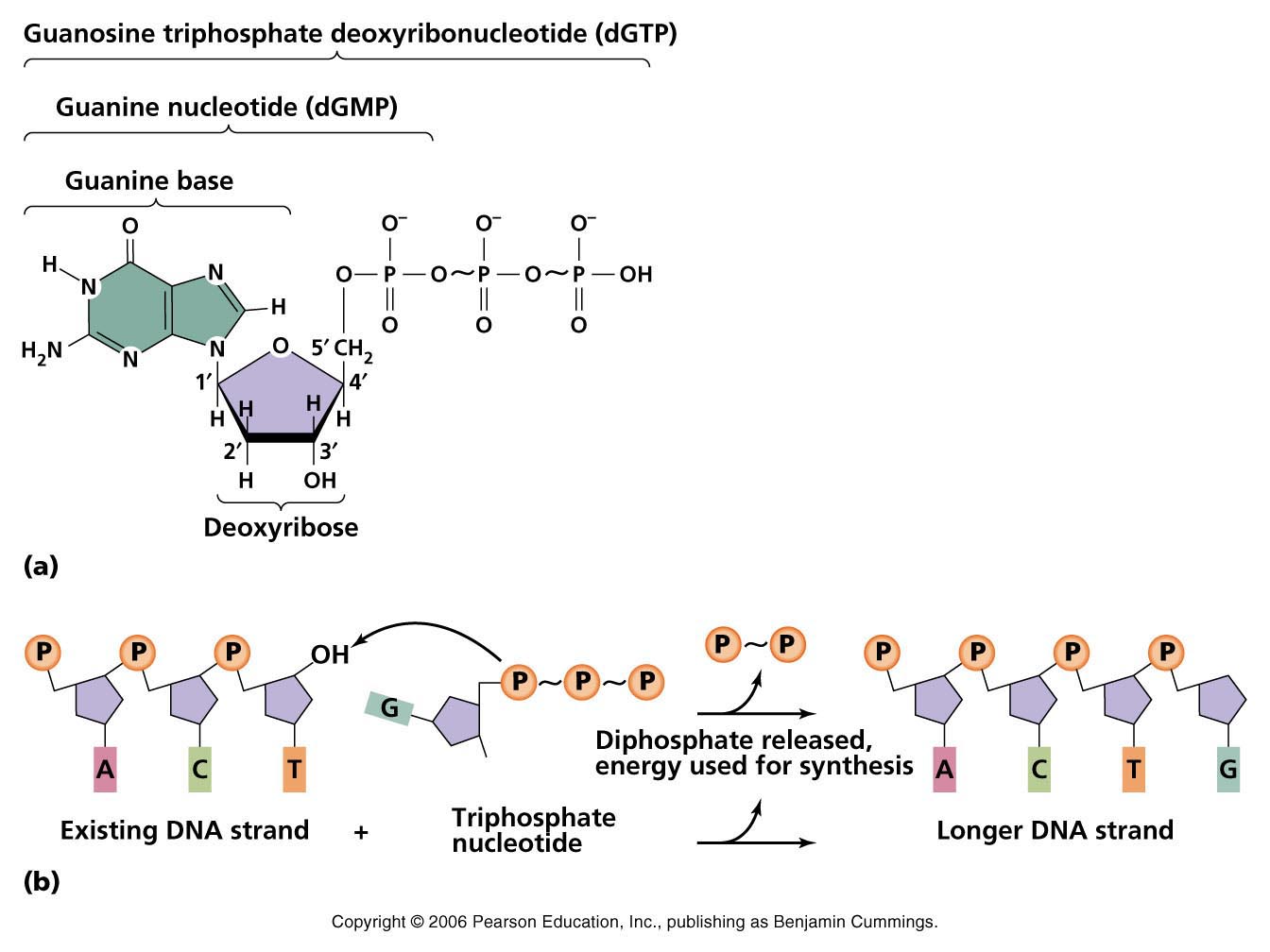
Все они передвигаются за счет гидролиза АТФ(разрушения молекулы с присоединением к получающимся в результате распада меньшим молекулам атомов, взятых из молекулы воды. Гидролиз показан на правой части схемы взаимопревращения АТФ и АДФ). Или за счет гидролиза ГТФ , отличающегося от АТФ только тем, что в его состав входит другой нуклеотид (гуанин).
Сценарий 3 : отщепление от АТФ или другой подобной молекулы, содержащей нуклеотид, сразу двух фосфатных групп приводит к еще большему выбросу энергии, чем когда отщепляется только один фосфат. Такой мощный выброс позволяет создавать прочный сахарофосфатный остов молекул ДНК и РНК:

1. для того, чтобы нуклеотиды могли присоединяться к строящейся цепи ДНК или РНК, их нужно активировать, присоединив две молекулы фосфата. Это энергозатратная реакция, выполняемая клеточными ферментами.
2. фермент ДНК- или РНК-полимераза (на схеме внизу не показан) присоединяет активированный нуклеотид (на схеме показан ГТФ) к строящемуся полинуклеотиду и катализирует отщепление двух фосфатных групп. Выделившаяся энергия используется на создание связи между фосфатной группой одного нуклеотида и рибозой другого. Созданные в результате связи не являются высокоэнергетичными, а значит разрушить их не просто, что является преимуществом для построения молекулы, содержащей наследственную информацию клетки или передающей ее.
В природе возможно спонтанное протекание только энергетически выгодных реакций, что обусловлено вторым законом термодинамики
Тем не менее живые клетки могут совмещать две реакции, одна из которых дает чуть больше энергии, чем поглощает вторая, и таким образом осуществлять энергозатратные реакции. Энергозатратные реакции направлены на создание из отдельных молекул и атомов более крупных молекул, клеточных органелл и целых клеток, тканей, органов и многоклеточных живых существ, а так же на запасание энергии для их метаболизма
Запасание энергии осуществляется за счет контролируемого и постепенного разрушения органических молекул (энергодающий процесс), сопряженного с созданием молекул-энергоносителей (энергозатратный процесс). Фотосинтезирующие организмы запасают таким образом энергию улавливаемых хлорофиллом солнечных фотонов
Молекулы-энергоносители делятся на две группы: хранящие энергию в виде высокоэнергетической связи или в виде присоединенного высокоэнергетического электрона. Впрочем, в первой группе высокая энергия обеспечивается таким же высокоэнергетическим электроном, так что можно сказать, что энергия запасается в загнанных на высокий уровень электронах, находящихся в составе разных молекул
Запасенная таким образом энергия отдается так же двумя способами: разрушением высокоэнергетической связи или передачей высокоэнергетических электронов для постепенного снижения их энергии. В обоих случаях энергия выделяется в виде испускания переходящим на более низкий энергетический уровень электроном частицы-переносчика электромагнитного поля (фотона) и тепла. Этот фотон улавливается таким образом, чтобы была совершена полезная работа (образование нужной для метаболизма молекулы в первом случае и прокачки протонов через мембрану митохондрии во втором)
Запасенная в виде протонного градиента энергия используется для синтеза АТФ, а также для других клеточных процессов, которые остались за рамками этой главы (думаю, никто не в обиде, учитывая ее размер). А синтезированная АТФ используется так, как описано в предыдущем пункте.
Продолжение. См. № 11, 12, 13, 14, 15, 16/2005
Уроки биологии в классах естественно-научного профиля
Расширенное планирование, 10 класс
Урок 19. Химическое строение и биологическая роль АТФ
Оборудование: таблицы по общей биологии, схема строения молекулы АТФ, схема взаимосвязи пластического и энергетического обменов.
I. Проверка знаний
Проведение биологического диктанта «Органические соединения живой материи»
Учитель читает тезисы под номерами, учащиеся записывают в тетрадь номера тех тезисов, которые подходят по содержанию их варианту.
Вариант 1 – белки.
Вариант 2 – углеводы.
Вариант 3 – липиды.
Вариант 4 – нуклеиновые кислоты.
1. В чистом виде состоят только из атомов С, Н, О.
2. Кроме атомов С, Н, О содержат атомы N и обычно S.
3. Кроме атомов С, Н, О содержат атомы N и Р.
4. Обладают относительно небольшой молекулярной массой.
5. Молекулярная масса может быть от тысяч до нескольких десятков и сотен тысяч дальтон.
6. Наиболее крупные органические соединения с молекулярной массой до нескольких десятков и сотен миллионов дальтон.
7. Обладают различными молекулярными массами – от очень небольшой до весьма высокой, в зависимости от того, является ли вещество мономером или полимером.
8. Состоят из моносахаридов.
9. Состоят из аминокислот.
10. Состоят из нуклеотидов.
11. Являются сложными эфирами высших жирных кислот.
12. Основная структурная единица: «азотистое основание–пентоза–остаток фосфорной кислоты».
13. Основная структурная единица: «аминокислот».
14. Основная структурная единица: «моносахарид».
15. Основная структурная единица: «глицерин–жирная кислота».
16. Молекулы полимеров построены из одинаковых мономеров.
17. Молекулы полимеров построены из сходных, но не вполне одинаковых мономеров.
18. Не являются полимерами.
19. Выполняют почти исключительно энергетическую, строительную и запасающую функции, в некоторых случаях – защитную.
20. Помимо энергетической и строительной выполняют каталитическую, сигнальную, транспортную, двигательную и защитную функции;
21. Осуществляют хранение и передачу наследственных свойств клетки и организма.
Вариант 1
– 2; 5; 9; 13; 17; 20.
Вариант 2
– 1; 7; 8; 14; 16; 19.
Вариант 3
– 1; 4; 11; 15; 18; 19.
Вариант 4
– 3; 6; 10; 12; 17; 21.
II. Изучение нового материала
1. Строение аденозинтрифосфорной кислоты
Кроме белков, нуклеиновых кислот, жиров и углеводов в живом веществе синтезируется большое количество других органических соединений. Среди них важнуую роль в биоэнергетике клетки играет аденозинтрифосфорная кислота (АТФ). АТФ содержится во всех клетках растений и животных. В клетках чаще всего аденозинтрифосфорная кислота присутствует в виде солей, называемых аденозинтрифосфатами . Количество АТФ колеблется и в среднем составляет 0,04% (в клетке в среднем находится около 1 млрд молекул АТФ). Наибольшее количество АТФ содержится в скелетных мышцах (0,2–0,5%).
Молекула АТФ состоит из азотистого основания – аденина, пентозы – рибозы и трех остатков фосфорной кислоты, т.е. АТФ – особый адениловый нуклеотид. В отличие от других нуклеотидов АТФ содержит не один, а три остатка фосфорной кислоты. АТФ относится к макроэргическим веществам – веществам, содержащим в своих связях большое количество энергии.
Пространственная модель (А) и структурная формула (Б) молекулы АТФ
Из состава АТФ под действием ферментов АТФаз отщепляется остаток фосфорной кислоты. АТФ имеет устойчивую тенденцию к отделению своей концевой фосфатной группы:
АТФ 4– + Н 2 О ––> АДФ 3– + 30,5 кДж + Фн,
т.к. это приводит к исчезновению энергетически невыгодного электростатического отталкивания между соседними отрицательными зарядами. Образовавшийся фосфат стабилизируется за счет образования энергетически выгодных водородных связей с водой. Распределение заряда в системе АДФ + Фн становится более устойчивым, чем в АТФ. В результате этой реакции высвобождается 30,5 кДж (при разрыве обычной ковалентной связи высвобождается 12 кДж).
Для того, чтобы подчеркнуть высокую энергетическую «стоимость» фосфорно-кислородной связи в АТФ, ее принято обозначать знаком ~ и называть макроэнергетической связью. При отщеплении одной молекулы фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорная кислота), а если отщепляются две молекулы фосфорной кислоты, то АТФ переходит в АМФ (аденозинмонофосфорная кислота). Отщепление третьего фосфата сопровождается выделением всего 13,8 кДж, так что собственно макроэргических связей в молекуле АТФ только две.
2. Образование АТФ в клетке
Запас АТФ в клетке невелик. Например, в мышце запасов АТФ хватает на 20–30 сокращений. Но ведь мышца способна работать часами и производить тысячи сокращений. Поэтому наряду с распадом АТФ до АДФ в клетке должен непрерывно идти обратный синтез. Существует несколько путей синтеза АТФ в клетках. Познакомимся с ними.
1. Анаэробное фосфорилирование. Фосфорилированием называют процесс синтеза АТФ из АДФ и низкомолекулярного фосфата (Фн). В данном случае речь идет о бескислородных процессах окисления органических веществ (например, гликолиз – процесс бескислородного окисления глюкозы до пировиноградной кислоты). Примерно 40% выделяемой в ходе этих процессов энергии (около 200 кДж/моль глюкозы), расходуется на синтез АТФ, а остальная часть рассеивается в виде тепла:
С 6 Н 12 О 6 + 2АДФ + 2Фн ––> 2С 3 Н 4 O 3 + 2АТФ + 4Н.
2. Окислительное фосфорилирование – это процесс синтеза АТФ за счет энергии окисления органических веществ кислородом. Этот процесс был открыт в начале 1930-х гг. XX в. В.А. Энгельгардтом. Кислородные процессы окисления органических веществ протекают в митохондриях. Примерно 55% выделяющейся при этом энергии (около 2600 кДж/моль глюкозы) превращается в энергию химических связей АТФ, а 45% рассеивается в виде тепла.
Окислительное фосфорилирование значительно эффективнее анаэробных синтезов: если в процессе гликолиза при распаде молекулы глюкозы синтезируется всего 2 молекулы АТФ, то в ходе окислительного фосфорилирования образуется 36 молекул АТФ.
3. Фотофосфорилирование – процесс синтеза АТФ за счет энергии солнечного света. Этот путь синтеза АТФ характерен только для клеток, способных к фотосинтезу (зеленые растения, цианобактерии). Энергия квантов солнечного света используется фотосинтетиками в световую фазу фотосинтеза для синтеза АТФ.
3. Биологическое значение АТФ
АТФ находится в центре обменных процессов в клетке, являясь связующим звеном между реакциями биологического синтеза и распада. Роль АТФ в клетке можно сравнить с ролью аккумулятора, так как в ходе гидролиза АТФ выделяется энергия, необходимая для различных процессов жизнедеятельности («разрядка»), а в процессе фосфорилирования («зарядка») АТФ вновь аккумулирует в себе энергию.

За счет выделяющейся при гидролизе АТФ энергии происходят почти все процессы жизнедеятельности в клетке и организме: передача нервных импульсов, биосинтез веществ, мышечные сокращения, транспорт веществ и др.
III. Закрепление знаний
Решение биологических задач
Задача 1. При быстром беге мы часто дышим, происходит усиленное потоотделение. Объясните эти явления.
Задача 2. Почему на морозе замерзающие люди начинают притопывать и подпрыгивать?
Задача 3. В известном произведении И.Ильфа и Е.Петрова «Двенадцать стульев» среди многих полезных советов можно найти и такой: «Дышите глубже, вы взволнованы». Попробуйте обосновать этот совет с точки зрения происходящих в организме энергетических процессов.
IV. Домашнее задание
Начать подготовку к зачету и контрольной работе (продиктовать вопросы зачета – см. урок 21).
Урок 20. Обобщение знаний по разделу «Химическая организация жизни»
Оборудование: таблицы по общей биологии.
I. Обобщение знаний раздела
Работа учащихся с вопросами (индивидуально) с последующими проверкой и обсуждением
1. Приведите примеры органических соединений, в состав которых входят углерод, сера, фосфор, азот, железо, марганец.
2. Как по ионному составу можно отличить живую клетку от мертвой?
3. Какие вещества находятся в клетке в нерастворенном виде? В какие органы и ткани они входят?
4. Приведите примеры макроэлементов, входящих в активные центры ферментов.
5. Какие гормоны содержат микроэлементы?
6. Какова роль галогенов в организме человека?
7. Чем белки отличаются от искусственных полимеров?
8. Чем отличаются пептиды от белков?
9. Как называется белок, входящий в состав гемоглобина? Из скольких субъединиц он состоит?
10. Что такое рибонуклеаза? Сколько аминокислот входит в ее состав? Когда она была синтезирована искусственно?
11. Почему скорость химических реакций без ферментов мала?
12. Какие вещества транспортируются белками через клеточную мембрану?
13. Чем отличаются антитела от антигенов? Содержат ли вакцины антитела?
14. На какие вещества распадаются белки в организме? Сколько энергии выделяется при этом? Где и как обезвреживается аммиак?
15. Приведите пример пептидных гормонов: как они участвуют в регуляции клеточного метаболизма?
16. Какова структура сахара, с которым мы пьем чай? Какие еще три синонима этого вещества вы знаете?
17. Почему жир в молоке не собирается на поверхности, а находится в виде суспензии?
18. Какова масса ДНК в ядре соматической и половой клеток?
19. Какое количество АТФ используется человеком в сутки?
20. Из каких белков люди изготавливают одежду?
Первичная структура панкреатической рибонуклеазы (124 аминокислоты)
II. Домашнее задание.
Продолжить подготовку к зачету и контрольной работе по разделу «Химическая организация жизни».
Урок 21. Зачетный урок по разделу «Химическая организация жизни»
I. Проведение устного зачета по вопросам
1. Элементарный состав клетки.
2. Характеристика органогенных элементов.
3. Структура молекулы воды. Водородная связь и ее значение в «химии» жизни.
4. Свойства и биологические функции воды.
5. Гидрофильные и гидрофобные вещества.
6. Катионы и их биологическое значение.
7. Анионы и их биологическое значение.
8. Полимеры. Биологические полимеры. Отличия периодических и непериодических полимеров.
9. Свойства липидов, их биологические функции.
10. Группы углеводов, выделяемые по особенностям строения.
11. Биологические функции углеводов.
12. Элементарный состав белков. Аминокислоты. Образование пептидов.
13. Первичная, вторичная, третичная и четвертичная структуры белков.
14. Биологические функция белков.
15. Отличия ферментов от небиологических катализаторов.
16. Строение ферментов. Коферменты.
17. Механизм действия ферментов.
18. Нуклеиновые кислоты. Нуклеотиды и их строение. Образование полинуклеотидов.
19. Правила Э.Чаргаффа. Принцип комплементарности.
20. Образование двухцепочечной молекулы ДНК и ее спирализация.
21. Классы клеточной РНК и их функции.
22. Отличия ДНК и РНК.
23. Репликация ДНК. Транскрипция.
24. Строение и биологическая роль АТФ.
25. Образование АТФ в клетке.
II. Домашнее задание
Продолжить подготовку к контрольной работе по разделу «Химическая организация жизни».
Урок 22. Контрольный урок по разделу «Химическая организация жизни»
I. Проведение письменной контрольной работы
Вариант 1
1. Имеются три вида аминокислот – А, В, С. Сколько вариантов полипептидных цепей, состоящих из пяти аминокислот, можно построить. Укажите эти варианты. Будут ли эти полипептиды обладать одинаковыми свойствами? Почему?
2. Все живое в основном состоит из соединений углерода, а аналог углерода – кремний, содержание которого в земной коре в 300 раз больше, чем углерода, встречается лишь в очень немногих организмах. Объясните этот факт с точки зрения строения и свойств атомов этих элементов.
3. В одну клетку ввели молекулы АТФ, меченные радиоактивным 32Р по последнему, третьему остатку фосфорной кислоты, а в другую – молекулы АТФ, меченные 32Р по первому, ближайшему к рибозе остатку. Через 5 минут в обеих клетках померили содержание неорганического фосфат-иона, меченного 32Р. Где оно окажется значительно выше?
4. Исследования показали, что 34% общего числа нуклеотидов данной иРНК приходится на гуанин, 18% – на урацил, 28% – на цитозин и 20% – на аденин. Определите процентный состав азотистых оснований двухцепочечной ДНК, слепком с которой является указанная иРНК.
Вариант 2
1. Жиры составляют «первый резерв» в энергетическом обмене и используются, когда исчерпан резерв углеводов. Однако в скелетных мышцах при наличии глюкозы и жирных кислот в большей степени используются последние. Белки же в качестве источника энергии всегда используются лишь в крайнем случае, при голодании организма. Объясните эти факты.
2. Ионы тяжелых металлов (ртути, свинца и др.) и мышьяка легко связываются сульфидными группировками белков. Зная свойства сульфидов этих металлов, объясните, что произойдет с белком при соединении с этими металлами. Почему тяжелые металлы являются ядами для организма?
3. В реакции окисления вещества А в вещество В освобождается 60 кДж энергии. Сколько молекул АТФ может быть максимально синтезировано в этой реакции? Как будет израсходована остальная энергия?
4. Исследования показали, что 27% общего числа нуклеотидов данной иРНК приходится на гуанин, 15% – на урацил, 18% – на цитозин и 40% – на аденин. Определите процентный состав азотистых оснований двухцепочечной ДНК, слепком с которой является указанная иРНК.
Продолжение следует
Практическое занятие № 15.
Задание к занятию № 15.
Тема: ЭНЕРГЕТИЧЕСКИЙ ОБМЕН.
Актуальность темы.
Биологическое окисление – совокупность протекающих в каждой клетке ферментативных процессов, в результате которых молекулы углеводов, жиров и аминокислот расщепляются, в конечном счете, до углекислоты и воды, а освобождающаяся энергия запасается клеткой в виде аденозинтрифосфорной кислоты (АТФ) и затем используется в жизнедеятельности организма (биосинтез молекул, процесс деления клеток, сокращение мышц, активный транспорт, продукция тепла и др.). Врач должен знать о существовании гипоэнергетических состояний, при которых снижается синтез АТФ. При этом страдают все процессы жизнедеятельности, которые протекают с использованием энергии, запасенной в виде макроэргических связей АТФ. Наиболее распространенная причина гипоэнергетических состояний – гипоксия тканей , связанная со снижением концентрации кислорода в воздухе, нарушением работы сердечно-сосудистой и дыхательной систем, анемиями различного происхождения. Кроме того, причиной гипоэнергетических состояний могут быть гиповитаминозы , связанные с нарушением структурного и функционального состояния ферментных систем, участвующих в процессе биологического окисления, а также голодание , которое приводит к отсутствию субстратов тканевого дыхания. Кроме того, в процессе биологического окисления образуются активные формы кислорода, запускающие процессы перекисного окисления липидов биологических мембран. Необходимо знать механизмы защиты организма от данных форм (ферменты, лекарственные препараты, оказывающие мембраностабилизирующее действие – антиоксиданты).
Учебные и воспитательные цели:
Общая цель занятия: привить знания о протекании биологического окисления, в результате которого образуется до 70-8- % энергии в виде АТФ, а также об образовании активных форм кислорода и их повреждающего действия на организм.
Частные цели: уметь определять пероксидазу в хрене, картофеле; активность сукцинатдегидрогеназы мышц.
1. Входной контроль знаний:
1.1. Тесты.
1.2. Устный опрос.
2. Основные вопросы темы:
2.1. Понятие об обмене веществ. Анаболические и катаболические процессы и их взаимосвязь.
2.2. Макроэргические соединения. АТФ – универсальный аккумулятор и источник энергии в организме. Цикл АТФ-АДФ. Энергетический заряд клетки.
2.3. Этапы обмена веществ. Биологическое окисление (тканевое дыхание). Особенности биологического окисления.
2.4. Первичные акцепторы протонов водорода и электронов.
2.5. Организация дыхательной цепи. Переносчики в дыхательной цепи (ЦПЭ).
2.6. Окислительное фосфорилирование АДФ. Механизм сопряжения окисления и фосфорилирования. Коэффициент окислительного фосфорилирования (Р/О).
2.7. Дыхательный контроль. Разобщение дыхания (окисления) и фосфорилирования (свободное окисление).
2.8. Образование токсичных форм кислорода в ЦПЭ и обезвреживание перекиси водорода ферментом пероксидазой.
Лабораторно-практические работы.
3.1. Методика определения пероксидазы в хрене.
3.2. Методика определения пероксидазы в картофеле.
3.3. Определение активности сукцинатдегидрогеназы мышц и конкурентное торможение её активности.
Выходной контроль.
4.1. Тесты.
4.2. Ситуационные задачи.
5. Литература:
5.1. Материалы лекций.
5.2. Николаев А.Я. Биологическая химия.-М.: Высшая школа, 1989., С 199-212, 223-228.
5.3. Березов Т.Т., Коровкин Б.Ф. Биологическая химия. - М.: Медицина, 1990.С.224-225.
5.4. Кушманова О.Д., Ивченко Г.М. Руководство к практическим занятиям по биохимии.- М.: Медицина, 1983, раб. 38.
2. Основные вопросы темы.
2.1. Понятие об обмене веществ. Анаболические и катаболические процессы и их взаимосвязь .
Живые организмы находятся в постоянной и неразрывной связи с окружающей средой.
Эта связь осуществляется в процессе обмена веществ.
Обмен веществ (метаболизм)– совокупность всех реакций в организме.
Промежуточный обмен (внутриклеточный метаболизм) – включает 2 типа реакций: катаболизм и анаболизм.
Катаболизм – процесс расщепления органических веществ до конечных продуктов (СО 2 , Н 2 О и мочевины). В этот процесс включаются метаболиты, образующиеся как при пищеварении, так и при распаде структурно-функциональных компонентов клеток.
Процессы катаболизма в клетках организма сопровождаются потреблением кислорода, который необходим для реакций окисления. В результате реакций катаболизма происходит выделение энергии (экзергонические реакции), которая необходима организму для его жизнедеятельности.
Анаболизм – синтез сложных веществ из простых. В анаболических процессах используется энергия, освобождающаяся при катаболизме (эндергонические реакции).
Источниками энергии для организма являются белки, жиры и углеводы. Энергия, заключенная в химических связях этих соединений, в процессе фотосинтеза трансформировалась из солнечной энергии.
Макроэргические соединения. АТФ – универсальный аккумулятор и источник энергии в организме. Цикл АТФ-АДФ. Энергетический заряд клетки.
АТФ – является макроэргическим соединением, содержащим макроэргические связи; при гидролизе концевой фосфатной связи выделяется около 20 кдж/моль энергии.
К макроэргическим соединениям относятся ГТФ, ЦТФ, УТФ, креатинфосфат, карбамоилфосфат и др. Они используются в организме для синтеза АТФ. Например, ГТФ + АДФ à ГДФ + АТФ
Этот процесс называется субстратное фосфорилирование – экзоргонические реакции. В свою очередь все эти макроэргические соединения образуются при использовании свободной энергии концевой фосфатной группы АТФ. Наконец, энергия АТФ используется для совершения различных видов работ в организме:
Механической (мышечное сокращение);
Электрической (проведение нервного импульса);
Химической (синтез веществ);
Осмотической (активный транспорт веществ через мембрану) – эндергонические реакции.
Таким образом, АТФ- главный, непосредственно используемый донор энергии в организме. АТФ занимает центральное место между эндергоническими и экзергоническими реакциями.
В организме человека образуется количество АТФ, равное массе тела и за каждые 24 часа вся эта энергия разрушается. 1 молекула АТФ «живет» в клетке около минуты.
Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счет энергии окисления органических соединений. Цикл АТФ-АДФ – основной механизм обмена энергии в биологических системах, а АТФ – универсальная «энергетическая валюта».
Каждая клетка обладает электрическим зарядом, который равен
[АТФ] + ½[АДФ]
[АТФ] + [АДФ] + [АМФ]
Если заряд клетки равен 0,8-0,9, то в клетке весь адениловый фонд представлен в виде АТФ (клетка насыщена энергией и процесс синтеза АТФ не происходит).
По мере использования энергии, АТФ превращается в АДФ, заряд клетки становится равным 0, автоматически начинается синтез АТФ.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
- Введение
- 1.1 Химические свойства АТФ
- 1.2 Физические свойства АТФ
- 2.1
- 3.1 Роль в клетке
- 3.2 Роль в работе ферментов
- 3.4 Другие функции АТФ
- Заключение
- Библиографический список
Перечень условных обозначений
АТФ - аденозинтрифосфат
АДФ - аденозиндифосфат
АМФ - аденозинмонофосфат
РНК - рибонуклеиновая кислота
ДНК - дезоксирибонуклеиновая кислота
НАД - никотинамидадениндинуклеотид
ПВК - пировиноградная кислота
Г-6-Ф - фосфоглюкозоизомераза
Ф-6-Ф - фруктоза-6-фосфат
ТПФ - тиаминпирофосфат
ФАД - фениладениндинуклеотид
Фн - неограниченный фосфат
G - энтропия
РНР - рибонуклеотидредуктаза
Введение
Основным источником энергии для всех живых существ, населяющих нашу планету, служит энергия солнечного света, которую используют непосредственно только клетки зеленых растений, водорослей, зеленых и пурпурных бактерий. В этих клетках из углекислого газа и воды в процессе фотосинтеза образуются органические вещества (углеводы, жиры, белки, нуклеиновые кислоты и др.). Поедая растения, животные получают органические вещества в готовом виде. Энергия, запасенная в этих веществах, переходит вместе с ними в клетки гетеротрофных организмов.
В клетках животных организмов энергия органических соединений при их окислении превращается в энергию АТФ. (Углекислый газ и вода, выделяющиеся при этом, вновь используются автотрофными организмами для процессов фотосинтеза.) За счет энергии АТФ осуществляются все процессы жизнедеятельности: биосинтез органических соединений, движение, рост, деление клеток и др.
Тема по образованию и использованию АТФ в организме давно не нова, но редко, где встретишь полное рассмотрение того и другого в одном источнике и еще реже анализ сразу обоих этих процессов и в разных организмах.
В связи с этим, актуальностью нашей работы стало тщательное изучение образования и использования АТФ в живых организмах, т.к. данная тема не изучается на должном уровне в научно-популярной литературе.
Целью нашей работы было:
· изучение механизмов образования и путей использования АТФ в организме животных и человека.
Перед нами были поставлены задачи:
· Изучить химическую природу и свойства АТФ;
· Проанализировать пути образования АТФ в живых организмах;
· Рассмотреть пути использования АТФ в живых организмах;
· Рассмотреть значение АТФ для организма человека и животных.
Глава 1. Химическая природа и свойства АТФ
1.1 Химические свойства АТФ
Аденозинтрифосфат - нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. АТФ был открыт в 1929 году Карлом Ломанном, а в1941 году Фриц Липман показал, что АТФ является основным переносчиком энергии в клетке .
Систематическое наименование АТФ:
9-в-D- рибофуранозиладенин-5"-трифосфат , или
9-в-D- рибофуранозил-6-амино-пурин-5"-трифосфат .
Химически АТФ представляет собой трифосфорный эфир аденозина, который является производным аденина и рибозы.
Пуриновое азотистое основание - аденин - соединяется в-N-гликозидной связью с 1"-углеродом рибозы. К 5"-углероду рибозы последовательно присоединяются три молекулы фосфорной кислоты, обозначаемые соответственно буквами: б, в и г .
По строению АТФ сходна с адениновым нуклеотидом, входящим в состав РНК, только вместо одной фосфорной кислоты в состав АТФ входят три остатка фосфорной кислоты. Клетки не в состоянии содержать кислоты в заметных количествах, а только их соли. Поэтому фосфорная кислота входит в АТФ в виде остатка (вместо ОН-группы кислоты имеется отрицательно заряженный атом кислорода).
Под действием ферментов молекула АТФ легко подвергается гидролизу, то есть присоединяет молекулу воды и расщепляется с образованием аденозиндифосфорной кислоты (АДФ):
АТФ + Н2О АДФ + Н3РО4.
Отщепление еще одного остатка фосфорной кислоты превращает АДФ в аденозинмонофосфорную кислоту АМФ:
АДФ + Н2О АMФ + Н3РО4.
Эти реакции обратимы, то есть АМФ может переходить в АДФ и далее в АТФ, аккумулируя энергию. Разрушение обычной пептидной связи высвобождает лишь 12 кДж/моль энергии. А связи, которыми присоединены остатки фосфорной кислоты, высокоэнергетичны (их еще называют макроэргическими): при разрушении каждой из них выделяется 40 кДж/моль энергии. Поэтому АТФ играет в клетках центральную роль универсального биологического аккумулятора энергии. Молекулы АТФ синтезируются в митохондриях и хлоропластах (лишь незначительное их количество синтезируется в цитоплазме), а затем поступают к различным органоидам клетки, обеспечивая энергией все процессы жизнедеятельности.
За счет энергии АТФ происходит деление клетки, активный перенос веществ через клеточные мембраны, поддержание мембранного электрического потенциала в процессе передачи нервных импульсов, а также биосинтез высокомолекулярных соединений и физическая работа.
При усиленной нагрузке (например, в беге на короткие дистанции) мышцы работают исключительно за счет запаса АТФ. В клетках мышц этого запаса хватает на несколько десятков сокращений, а дальше количество АТФ должно восполняться. Синтез АТФ из АДФ и АМФ происходит за счет энергии, выделяющейся при расщеплении углеводов, липидов и других веществ. На выполнение умственной работы также затрачивается большое количество АТФ. По этой причине людям умственного труда требуется повышенное количество глюкозы, расщепление которой обеспечивает синтез АТФ .
1.2 Физические свойства АТФ
АТФ состоит из аденозина и рибозы - и трех фосфатных групп. ATФ хорошо растворим в воде и довольно устойчивы в растворах при рН 6.8-7.4, но быстро гидролизуется в экстремальных рН. Следовательно, АТФ лучше всего хранить в безводных солях.
АТФ является неустойчивой молекулой. В небуферизованной воде, он гидролизуется до АДФ и фосфата. Это потому, что прочность связей между фосфатными группами в АТФ меньше, чем прочность водородных связей (гидратации облигации), между своей продукции (АДФ + фосфат), и воду. Таким образом, если АТФ и АДФ находятся в химическом равновесии в воде, почти все АТФ будет в конечном счете превращается в AДФ. Система, которая далека от равновесия содержит свободную энергию Гиббса, и способен делать работу. Живые клетки поддерживают соотношение АТФ в АДФ в точке десять порядков от равновесия, при концентрации АТФ в тысячу раз выше, чем концентрация AДФ. Это смещение от положения равновесия означает, что гидролиз АТФ в клетке выпускает большое количество свободной энергии .
Две высокоэнергетические фосфатные связи (те, которые соединяют соседние фосфаты) в молекулы АТФ несут ответственность за высокое содержание энергии этой молекулы. Энергия, запасенная в АТФ может быть освобождена от гидролиза. Расположенный дальний от сахара рибозы, г-фосфатная группа имеет более высокую энергию гидролиза, чем любой б - или в-фосфат. Связи, образованные после гидролиза или фосфорилирования остатка АТФ - ниже по энергии, чем других связей АТФ. Во время фермент-катализируемого гидролиза АТФ или АТФ фосфорилирования, имеющиеся свободная энергия может быть использована на живые системы, чтобы сделать работу .
Любая нестабильная система потенциально реактивных молекул потенциально может служить способом хранения свободной энергии, если клетки сохранили свою концентрацию далеко от точки равновесия реакции. Однако, как и в случае с большинством полимерных биомолекул, разбивка РНК, ДНК и АТФ в простые мономеры обусловлена как выделение энергии и энтропии, увеличение соображения, как в стандартной концентрации, а также тех концентрациях, в которых встречается в клетке .
Стандартное количество энергии, выделяющейся в результате гидролиза АТФ может быть рассчитана по изменениям в энергии, не связанных с природными (стандартные) условия, то исправление биологической концентрации. Чистое изменение в тепловой энергии (энтальпии) при стандартной температуре и давлении разложения АТФ в АДФ и неорганических фосфатов - 20,5 кДж / моль, с изменением свободной энергии на 3,4 кДж / моль. Энергия выпускается путем расщепления фосфата или пирофосфата от АТФ на государственный стандарт 1 М являются:
АТФ + H 2 O > АДФ + P я ДG ? = - 30,5 кДж / моль (-7,3 ккал / моль)
АТФ + H 2 O > AMP + PP я ДG ? = - 45,6 кДж / моль (-10,9 ккал / моль)
Эти значения могут быть использованы для расчета изменения энергии в физиологических условиях и клеточных АТФ / АДФ. Тем не менее, более представительная значимость, называемая энергетическим зарядом, чаще работает. Значения приведены для свободной энергии Гиббса. Эти реакции зависят от ряда факторов, включая общую ионную силу и присутствие щелочно-земельных металлов, такие как ионы Mg 2 + и Ca 2 + . В обычных условиях, ДG составляет около - 57 кДж / моль (-14 ккал / моль) .
белок биологический аккумулятор энергия
Глава 2. Пути образования АТФ
В организме АТФ синтезируется путём фосфорилирования АДФ:
АДФ + H 3 PO 4 + энергия > АТФ + H 2 O.
Фосфорилирование АДФ возможно двумя способами: субстратное фосфорилирование и окислительное фосфорилирование (используя энергию окисляющихся веществ). Основная масса АТФ образуется на мембранах митохондрий в ходе окислительного фосфорилирования H-зависимой АТФ-синтазой. Субстратное фосфорилирование АТФ не требует участия мембранных ферментов, оно происходит в процессе гликолиза или путём переноса фосфатной группы с других макроэргических соединений.
Реакции фосфорилирования АДФ и последующего использования АТФ в качестве источника энергии образуют циклический процесс, составляющий суть энергетического обмена .
В организме АТФ является одним из самых часто обновляемых веществ. Так у человека продолжительность жизни одной молекулы АТФ менее 1 мин. В течение суток одна молекула АТФ проходит в среднем 2000-3000 циклов ресинтеза (человеческий организм синтезирует около 40 кг АТФ в день), то есть запаса АТФ в организме практически не создаётся, и для нормальной жизнедеятельности необходимо постоянно синтезировать новые молекулы АТФ .
Окислительное фосфорилирование -
Однако чаще всего в качестве субстрата используются углеводы. Так, клетки головного мозга не способны использовать для питания никакой другой субстрат, кроме углеводов.
Предварительно сложные углеводы расщепляются до простых, вплоть до образования глюкозы. Глюкоза является универсальным субстратом в процессе клеточного дыхания. Окисление глюкозы подразделяется на 3 этапа:
1. гликолиз;
2. окислительное декарбоксилирование и цикл Кребса;
3. окислительное фосфорилирование.
При этом гликолиз является общей фазой для аэробного и анаэробного дыхания .
2 .1.1 Гл ико лиз - ферментативный процесс последовательного расщепления глюкозы в клетках, сопровождающийся синтезом АТФ. Гликолиз при аэробных условиях ведёт к образованию пировиноградной кислоты (пирувата), гликолиз в анаэробных условиях ведёт к образованию молочной кислоты (лактата). Гликолиз является основным путём катаболизма глюкозы в организме животных .
Гликолитический путь представляет собой 10 последовательных реакций, каждая из которых катализируется отдельным ферментом.
Процесс гликолиза условно можно разделить на два этапа. Первый этап, протекающий с расходом энергии 2-х молекул АТФ, заключается в расщеплении молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся синтезом АТФ. Сам по себе гликолиз является полностью анаэробным процессом, то есть не требует для протекания реакций присутствия кислорода.
Гликолиз - один из древнейших метаболических процессов, известный почти у всех живых организмов. Предположительно гликолиз появился более 3,5 млрд. лет назад у первичных прокариот .
Результатом гликолиза является превращение одной молекулы глюкозы в две молекулы пировиноградной кислоты (ПВК) и образование двух восстановительных эквивалентов в виде кофермента НАД H.
Полное уравнение гликолиза имеет вид:
С 6 Н 12 О 6 + 2НАД + + 2АДФ + 2Ф н = 2НАД Н + 2ПВК + 2АТФ + 2H 2 O + 2Н + .
При отсутствии или недостатке в клетке кислорода пировиноградная кислота подвергается восстановлению до молочной кислоты, тогда общее уравнение гликолиза будет таким:
С 6 Н 12 О 6 + 2АДФ + 2Ф н = 2лактат + 2АТФ + 2H 2 O.
Таким образом, при анаэробном расщеплении одной молекулы глюкозы суммарный чистый выход АТФ составляет две молекулы, полученные в реакциях субстратного фосфорилирования АДФ.
У аэробных организмов конечные продукты гликолиза подвергаются дальнейшим превращениям в биохимических циклах, относящихся к клеточному дыханию. В итоге после полного окисления всех метаболитов одной молекулы глюкозы на последнем этапе клеточного дыхания - окислительном фосфорилировании, происходящем на митохондриальной дыхательной цепи в присутствии кислорода, - дополнительно синтезируются ещё 34 или 36 молекулы АТФ на каждую молекулу глюкозы.
Первой реакцией гликолиза является фосфорилирование молекулы глюкозы, происходящее при участии тканеспецифичного фермента гексокиназы с затратой энергии 1 молекулы АТФ; образуется активная форма глюкозы - глюкозо-6-фосфат (Г-6-Ф ):
Для протекания реакции необходимо наличие в среде ионов Mg 2+ , с которым комплексно связывается молекула АТФ. Эта реакция необратима и является первой ключевой реакцией гликолиза .
Фосфорилирование глюкозы преследует две цели: во-первых, из-за того что плазматическая мембрана, проницаемая для нейтральной молекулы глюкозы, не пропускает отрицательно заряженные молекулы Г-6-Ф, фосфорилированная глюкоза оказывается запертой внутри клетки. Во-вторых, при фосфорилировании глюкоза переводится в активную форму, способную участвовать в биохимических реакциях и включаться в метаболические циклы.
Печёночный изофермент гексокиназы - глюкокиназа - имеет важное значение в регуляции уровня глюкозы в крови.
В следующей реакции (2 ) ферментом фосфоглюкоизомеразой Г-6-Ф превращается во фруктозо-6-фосфат (Ф-6-Ф ):
Энергия для этой реакции не требуется, и реакция является полностью обратимой. На данном этапе в процесс гликолиза может также включаться путём фосфорилирования и фруктоза.
Далее почти сразу друг за другом следуют две реакции: необратимое фосфорилирование фруктозо-6-фосфата (3 ) и обратимое альдольное расщепление образовавшегося фруктозо-1,6-бифосфата (Ф-1,6-бФ ) на две триозы (4 ).
Фосфорилирование Ф-6-Ф осуществляется фосфофруктокиназой с затратой энергии ещё одной молекулы АТФ; это вторая ключевая реакция гликолиза, её регуляция определяет интенсивность гликолиза в целом.
Альдольное расщепление Ф-1,6-бФ происходит под действием альдолазы фруктозо-1,6-бифосфата:
В результате четвёртой реакции образуются дигидроксиацетонфосфат и глицеральдегид-3-фосфат , причём первый почти сразу под действием фосфотриозоизомеразы переходит во второй (5 ), который и участвует в дальнейших превращениях:
Каждая молекула глицеральдегидфосфата окисляется НАД + в присутствии дегидрогеназы глицеральдегидфосфата до 1,3- д ифосфоглице - рата (6 ):
Далее с 1,3-дифосфоглицерата , содержащего макроэргическую связь в 1 положении, ферментом фосфоглицераткиназой на молекулу АДФ переносится остаток фосфорной кислоты (реакция 7 ) - образуется молекула АТФ:
Это первая реакция субстратного фосфорилирования. С этого момента процесс расщепления глюкозы перестаёт быть убыточным в энергетическом плане, так как энергетические затраты первого этапа оказываются компенсированными: синтезируются 2 молекулы АТФ (по одной на каждый 1,3-дифосфоглицерат) вместо двух потраченных в реакциях 1 и 3 . Для протекания данной реакции требуется присутствие в цитозоле АДФ, то есть при избытке в клетке АТФ (и недостатке АДФ) её скорость снижается. Поскольку АТФ, не подвергающийся метаболизму, в клетке не депонируется а просто разрушается, то эта реакция является важным регулятором гликолиза.
Затем последовательно: фосфоглицеролмутаза образует 2-фосфо - глицерат (8 ):
Енолаза образует фосфоенолпируват (9 ):
И наконец происходит вторая реакция субстратного фосфорилирования АДФ с образованием енольной формы пирувата и АТФ (10 ):
Реакция протекает под действием пируваткиназы. Это последняя ключевая реакция гликолиза. Изомеризация енольной формы пирувата в пируват происходит неферментативно.
С момента образования Ф-1,6-бФ с выделением энергии протекают только реакции 7 и 10 , в которых и происходит к субстратное фосфорилирование АДФ .
Регуляция гликолиза
Различают местную и общую регуляцию.
Местная регуляция осуществляется путём изменения активности ферментов под действием различных метаболитов внутри клетки.
Регуляция гликолиза в целом, сразу для всего организма, происходит под действием гормонов, которые, влияя через молекулы вторичных посредников, изменяют внутриклеточный метаболизм.
Важное значение в стимуляции гликолиза принадлежит инсулину. Глюкагон и адреналин являются наиболее значимыми гормональными ингибиторами гликолиза.
Инсулин стимулирует гликолиз через:
· активацию гексокиназной реакции;
· стимуляцию фосфофруктокиназы;
· стимуляцию пируваткиназы.
Также на гликолиз влияют и другие гормоны. Например, соматотропин ингибирует ферменты гликолиза, а тиреоидные гормоныявляются стимуляторами.
Регуляция гликолиза осуществляется через несколько ключевых этапов. Реакции, катализируемые гексокиназой (1 ), фосфофруктокиназой (3 ) и пируваткиназой (10 ) отличаются существенным уменьшением свободной энергии и являются практически необратимыми, что позволяет им быть эффективными точками регуляции гликолиза .
Гликолиз - катаболический путь исключительной важности. Он обеспечивает энергией клеточные реакции, в том числе и синтез белка. Промежуточные продукты гликолиза используются при синтезе жиров. Пируват также может быть использован для синтезааланина, аспартата и других соединений. Благодаря гликолизу производительность митохондрий и доступность кислорода не ограничивают мощность мышц при кратковременных предельных нагрузках .
2.1.2 Окислительное декарбоксилирование - окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название "пируватдегидрогеназный комплекс" .
На I стадии этого процесса пируват теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E 1). На II стадии оксиэтильная группа комплекса E 1 -ТПФ-СНОН-СН 3 окисляется с образованием ацетильнойгруппы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментомдигидроли-поилацетилтрансферазой (Е 2). Этот фермент катализирует III стадию - перенос ацетильнойгруппы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид-Е 2 . При участии фермента дигидролипоилдегидрогеназы (Е 3) осуществляется переносатомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН 2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н + .
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E 1 , ли-поамид-Е 2 и ФАД-Е 3), а два - легко диссоциируют (HS-KoA и НАД).
Рис. 1 Механизм действия пируватдегидрогеназного комплекса
Е 1 - пируватдегидрогеназа; Е 2 - ди-гидролипоилацетилтрансфсраза; Е 3 - дигидролипоилдегидрогеназа; цифры в кружках обозначают стадии процесса.
Все эти ферменты, имеющие субъединичное строение, и коферменты организованы в единый комплекс. Поэтому промежуточные продукты способны быстро взаимодействовать друг с другом. Показано, что составляющие комплекс полипептидные цепи субъединиц дигидролипоил-ацетилтрансферазы составляют как бы ядро комплекса, вокруг которого расположены пируватдегидрогеназа и дигидролипоилдегидрогеназа. Принято считать, что нативный ферментный комплекс образуется путем самосборки.
Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом:
Пируват + НАД + + HS-KoA - > Ацетил-КоА + НАДН + Н + + СO 2 .
Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима.
Образовавшийся в процессе окислительного декарбоксилирования аце-тил-КоА подвергается дальнейшемуокислению с образованием СО 2 и Н 2 О. Полное окисление ацетил-КоА происходит в цикле трикарбоновых кислот (цикл Кребса). Этот процесс, так же как окислительное декарбо-ксилирование пирувата, происходит вмитохондриях клеток .
2 .1.3 Цикл трикарбоновых кисло т (цикл Кре бса , цитра тный цикл ) - центральная часть общего пути катаболизма, циклический биохимический аэробный процесс, в ходе которого происходит превращение двух - и трёхуглеродных соединений, образующихся как промежуточные продукты в живых организмах при распаде углеводов, жиров и белков, до CO 2 . При этом освобождённый водород направляется в цепь тканевого дыхания, где в дальнейшем окисляется до воды, принимая непосредственное участие в синтезе универсального источника энергии - АТФ.
Цикл Кребса - это ключевой этап дыхания всех клеток, использующих кислород, центр пересечения множества метаболических путей в организме. Кроме значительной энергетической роли циклу отводится также и существенная пластическая функция, то есть это важный источник молекул-предшественников, из которых в ходе других биохимических превращений синтезируются такие важные для жизнедеятельности клетки соединения как аминокислоты, углеводы, жирные кислоты и др.
Цикл превращения лимонной кислоты в живых клетках был открыт и изучен немецким биохимиком сэром Хансом Кребсом, за эту работу он (совместно с Ф. Липманом) был удостоен Нобелевской премии (1953 год).
У эукариот все реакции цикла Кребса протекают внутри митохондрий, причём катализирующие их ферменты, кроме одного, находятся в свободном состоянии в митохондриальном матриксе, исключение составляет сукцинатдегидрогеназа, которая локализуется на внутренней митохондриальной мембране, встраиваясь в липидный бислой. У прокариот реакции цикла протекают в цитоплазме .
Общее уравнение одного оборота цикла Кребса:
Ацетил-КоА > 2CO 2 + КоА + 8e ?
Регуляция цикл а :
Цикл Кребса регулируется "по механизму отрицательной обратной связи", при наличии большого количества субстратов (ацетил-КоА, оксалоацетат), цикл активно работает, а при избытке продуктов реакции (НАД, ATФ) тормозится. Регуляция осуществляется и при помощи гормонов, основным источником ацетил-КоА является глюкоза, поэтому гормоны, способствующие аэробному распаду глюкозы, способствуют работе цикла Кребса. Такими гормонами являются:
· инсулин;
· адреналин.
Глюкагон стимулирует синтез глюкозы и ингибирует реакции цикла Кребса.
Как правило работа цикла Кребса не прерывается за счёт анаплеротических реакций, которые пополняют цикл субстратами:
Пируват + СО 2 + АТФ = Оксалоацетат (субстрат Цикла Кребса) + АДФ + Фн.
Работа АТФ-синтазы
Процесс окислительного фосфорилирования осуществляется пятым комплексом дыхательной цепи митохондрий - Протонная АТФ-синтаза, состоящая из 9 субъединиц 5 типов:
· 3 субъединицы (г,д,е) способствуют целостности АТФ-синтазы
· в субъединица является основной функциональной единицей. Она имеет 3 конформации:
· L-конформация - присоединяет АДФ и Фосфат (поступают в митохондрию из цитоплазмы с помощью специальных переносчиков)
· Т-конформация - к АДФ присоединяется фосфат и образуется АТФ
· О-конформация - АТФ отщепляется от в-субъединицы и переходит на б-субъединицу.
· Для того, чтобы субъединица изменила конформацию необходим протон водорода, так как конформация меняется 3 раза необходимо 3 протона водорода. Протоны перекачиваются из межмембранного пространства митохондрии под действием электрохимического потенциала.
· б-субъединица транспортирует АТФ к мембранному переносчику, который "выбрасывает" АТФ в цитоплазму. Взамен из цитоплазмы этот же переносчик транспортирует АДФ. На внутренней мембране митохондрий также находится переносчик Фосфата из цитоплазмы в митохондрию, но для его работы необходим протон водорода. Такие переносчики называются транслоказами .
Суммарный выход
Для синтеза 1 молекулы АТФ необходимо 3 протона.
Ингибиторы окислительного фосфорилирования
Ингибиторы блокируют V комплекс:
· Олигомицин - блокируют протонные каналы АТФ-синтазы.
· Атрактилозид, циклофиллин - блокируют транслоказы.
Разобщители окислительного фосфорилирования
Разобщители - липофильные вещества, которые способны принимать протоны и переносить их через внутреннюю мембрану митохондрий минуя V комплекс (его протонный канал). Разобщители:
· Естественные - продукты перекисного окисления липидов, жирных кислот с длинной цепью; большие дозы тиреоидных гормонов.
· Искусственные - динитрофенол, эфир, производные витамина К, анестетики .
2.2 Субстратное фосфорилирование
Субстр атное фосфорил ирование (биохимическое), синтез богатых энергией фосфорных соединений за счёт энергии окислительно-восстановительных реакций гликолиза (катализируемых фосфоглице-ральдегиддегидрогеназой и енолазой) и при окислении a-кетоглутаровой кислоты в цикле трикарбоновых кислот (под действием a-кетоглутаратдегидрогеназы и сукцинаттиокиназы). Для бактерий описаны случаи С. ф. при окислении пировиноградной кислоты.С. ф., в отличие от фосфорилирования в цепи переноса электронов, не ингибируется "разобщающими" ядами (например, динитрофенолом) и не связано с фиксацией ферментов в мембранах митохондрий. Вклад С. ф. в клеточный фонд АТФ в аэробных условиях значительно меньше, чем вклад фосфорилирования в цепи переноса электронов .
Глава 3. Пути использования АТФ
3.1 Роль в клетке
Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. Всё это реакции синтеза сложных веществ в организме: осуществление активного переноса молекул через биологические мембраны, в том числе и для создания трансмембранного электрического потенциала; осуществления мышечного сокращения .
Как известно в биоэнергетике живых организмов имеют значение два основных момента:
а) химическая энергия запасается путём образования АТФ, сопряжённого с экзергоническими катаболическими реакциями окисления органических субстратов;
б) химическая энергия утилизируется путём расщепления АТФ, сопряжённого с эндергоническими реакциями анаболизма и другими процессами, требующими затраты энергии .
Встаёт вопрос, почему молекула АТФ соответствует своей центральной роли в биоэнергетике. Для его разрешения рассмотрим структуру АТФ Структура АТФ - (при рН 7,0 тетразаряд аниона ) .
АТФ представляет собой термодинамически нестойкое соединение. Нестабильность АТФ определяется, во - первых, электростатическим отталкиванием в области кластера одноимённых отрицательных зарядов, что приводит к напряжению всей молекулы, однако сильнее всего связи - Р - О - Р, и во - вторых, конкретным резонансом. В соответствии с последним фактором существует конкуренция между атомами фосфора за неподелённые подвижные электроны атома кислорода, расположенного между ними, поскольку на каждом атоме фосфора имеется частичный положительный заряд в следствии значительного электронаицепторного влияния групп Р=О и Р - О-. Таким образом, возможность существования АТФ определяется наличием достаточного количества химической энергии в молекуле, позволяющей компенсировать эти физико - химические напряжения. В молекуле АТФ имеется две фосфоангидридных (пирофосфатных) связи, гидролиз которых сопровождается значительным уменьшением свободной энергии (при рН 7,0 и 37 о С).
АТФ+Н 2 О = АДФ + Н 3 РО 4 G0I = - 31,0 КДж/моль.
АДФ+Н 2 О = АМФ +Н 3 РО 4 G0I = - 31,9 КДж/моль.
Одной из центральных проблем биоэнергетики является биосинтез АТФ, который в живой природе происходит путём Фосфорилирование АДФ.
Фосфорилирование АДФ является эндергоническим процессом и требует источника энергии. Как отмечалось ранее, в природе преобладает два таких источника энергии - это солнечная энергия и химическая энергия восстановленных органических соединений. Зелёные растения и некоторые микроорганизмы способны трансформировать энергию, поглощённых квантов света в химическую энергию, которая расходуется на фосфорилирование АДФ в световой стадии фотосинтеза. Этот процесс регенерации АТФ получил название фотосинтетического фосфорилирования. Трансформация энергии окисления органических соединений в макроэнергетические связи АТФ в аэробных условиях происходит преимущественно путём окислительного фосфорилирования. Свободная энергия, необходимая для образования АТФ, генерируется в дыхательной окислительной цепи митаходрий.
Известен ещё один тип синтеза АТФ, получивший название субстратного фосфорилирования. В отличии от окислительного фосфорилирования, сопряжённого с переносом электронов, донором активированной фосфорильной группой (- РО3 Н2), необходимой для регенерации АТФ, являются интермедианты процессов гликолиза и цикла трикарбоновых кислот. Во всех этих случаях окислительные процессы приводят к образованию высокоэнергетических соединений: 1,3 - дифосфоглицерата (гликолиз), сукцинил - КоА (цикл трикарбоновых кислот), которые при участии соответствующих ферментов способны фолирировать АДФ и образовывать АТФ. Трансформация энергии на уровне субстрата является единственным путём синтеза АТФ в анаэробных организмах. Этот процесс синтеза АТФ позволяет поддерживать интенсивную работу скелетных мышц в периоды кислородного голодания. Следует помнить, что он является единственным путём синтеза АТФ в зрелых эритроцитах не имеющих митохондрий.
Особо важную роль в биоэнергетике клетки играет адениловый нуклеотид, и которому присоединены два остатка фосфорной кислоты. Такой вещество называется аденозинтрифосфорной кислотой (АТФ). В химических связях между остатками фосфорной кислоты молекулы АТФ запасена энергия, которая освобождается при отщеплении органического фосфорита:
АТФ= АДФ+Ф+Е,
где Ф - фермент, Е - освобождающая энергия. В этой реакции образуется аденозинфосфорная кислота (АДФ) - остаток молекулы АТФ и органический фосфат. Энергию АТФ все клетки используют для процессов биосинтеза, движения, производство тепла, нервных импульсов, свечений (например, улюминисцентных бактерий), то есть для всех процессов жизнедеятельности .
АТФ - универсальный биологический аккумулятор энергии. Световая энергия, заключенная в потребляемой пище, запасается в молекулы АТФ.
Запас АТФ в клетке невелик. Так, в мышце запаса АТФ хватает на 20 - 30 сокращений. При усиленной, но кратковременной работе мышцы работают исключительно за счёт расщепления содержащейся в них АТФ. После окончания работы человек усиленно дышит - в этот период происходит расщепление углеводов и других веществ (происходит накопление энергии) и запас АТФ в клетках восстанавливается.
Также известна роль АТФ в качестве медиатора в синапсах .
3.2 Роль в работе ферментов
Живая клетка является далека от равновесия химическая система: ведь приближение живой системы к равновесию означает ее распад и смерть. Продукт каждого фермента обычно быстро расходуется, поскольку используется в качестве субстрата другим ферментом данного метаболического пути. Еще более важно, что большое количество ферментативных реакций связана с расщеплением АТФ на АДФ и неорганический фосфат. Чтобы это было возможным, пул АТФ в свою очередь должен поддерживаться на уровне, далеком от равновесия, так чтобы отношение концентрации АТФ в концентрации продуктов его гидролиза было высоким. Таким образом, пул АТФ играет роль "аккумулятора", поддерживающий постоянный перенес в клетке энергии и атомов по метаболическим путям, определяются присутствовать ферментами.
Итак, рассмотрим процесс гидролиза АТФ и его влияние на работу ферментов. Представим себе типичный биосинтетических процессов, при котором два мономеры - А и Б - должны объединиться между собой в реакции дегидратации (ее также называют конденсацией), сопровождающееся выделением воды:
А - Н + Б - ОН - АБ + Н2О
Обратная реакция, которую называют гидролизом, в которой молекула воды разрушает ковалентно связанную соединение А - Б, почти всегда будет энергетически выгодной. Это имеет место, например, при гидролитическом расщеплении белков, нуклеиновых кислот и полисахаридов на субъединицы.
Общая стратегия, по которой происходит образование клетки А - Б с А - Н и Б - ОН, включает в себя многоступенчатую последовательность реакций, в результате которых происходит н Связывание энергетически невыгодного синтеза нужных соединений со сбалансированной выгодной реакцией.
Гдролизу АТФ соответствует большая отрицательная величина? G, поэтому гидролиз АТФ часто играет роль энергетически благоприятной реакции, благодаря которой осуществляются внутриклеточные реакции биосинтеза.
На пути от А - Н и Б - ОН-А - Б, связанном с гидролизом АТФ, энергия гидролиза сначала переводит Б - ОН в высокоэнергетическую промежуточную соединение, которое затем непосредственно реагирует с А - Н, образуя А - Б. простой механизм данного процесса включает в себя перенес фосфата от АТФ к Б - ОН с образованием Б - ОРО 3 , или Б - О - Р, причем в этом случае суммарная реакция происходит лишь в две стадии:
1) Б - ОН + АТФ - Б - В - Р + АДФ
2) А - Н + Б - О - Р - А - Б + Р
Поскольку промежуточное соединение Б - О - Р, образующегося в процессе реакции, вновь разрушается, суммарные реакции можно описать с помощью следующих уравнений:
3) А-Н + Б - ОН - А - Б и АТФ - АДФ + Р
Первая, энергетически невыгодна реакция, оказывается возможной потому, что она связана со второй, энергетически выгодной реакцией (гидролиз АТФ). Примером связанных биосинтетических реакций подобного типа может быть синтез аминокислоты глутамина.
Величина G гидролиза АТФ до АДФ и неорганического фосфата зависит от концентрации всех реагирующих веществ и обычно для клетки условиях лежит в пределах от - 11 до - 13 ккал / моль. Реакция гидролиза АТФ, наконец, может быть использована для осуществления термодинамически невыгодной реакции со значением G, равным примерно +10 ккал / моль, конечно, в присутствии соответствующей последовательности реакций. Однако для многих реакций биосинтеза оказывается недостаточным даже? G = - 13 ккал / моль. В этих и других случаях путь гидролиза АТФ изменяется таким образом, что сначала образуются АМФ и РР (пирофосфат). На следующей стадии пирофосфат также подвергается гидролизу; общее изменение свободной энергии всего процесса составляет примерно - 26 ккал / моль.
Каким образом энергия гидролиза пирофосфата используется в биосинтетических реакциях? Один из путей можно продемонстрировать на примере приведенного выше синтеза соединения А - Б с А - Н и Б - ОН. При помощи соответствующего фермента Б - ОН может вступить в реакцию с АТФ и превратиться в высокоэнергетическую соединение Б - О - Р - Р. Теперь реакция состоит из трех стадий:
1) Б - ОН + АТФ - Б - В - Р - Р + АМФ
2) А - Н + Б - О - Р - Р - А - Б + РР
3) РР + Н2О - 2Р
Суммарную реакцию можно представить в следующем виде:
А - Н + Б - ОН - А - Б и АТФ + Н2О - АМФ + 2Р
Поскольку фермент всегда ускоряет катализовану ним реакцию как в прямом, так и в обратном направлении, соединение А - Б может распадаться, реагируя с пирофосфатом (реакция, обратная стадии 2). Однако энергетически выгодная реакция гидролиза пирофосфата (стадия 3) способствует поддержанию стабильности соединения А-Б за счет того, что концентрация пирофосфата остается очень низкой (это предотвращает протекание реакции, обратной к стадии 2). Таким образом, энергия гидролиза пирофосфата обеспечивает протекание реакции в прямом направлении. Примером важной биосинтетических реакции такого типа является синтез полинуклеотидов .
3.3 Роль в синтезе ДНК и РНК и белков
Во всех известных организмах, дезоксирибонуклеотиды, которые составляют ДНК синтезируется под действием ферментов рибонуклеотид-редуктазы (РНР) на соответствующих рибонуклеотидах. Эти ферменты снижают остатка сахара отрибозы, чтобы дезоксирибоза путем удаления кислорода из 2" гидроксильные группы, субстраты рибонуклеозид дифосфаты и продукты дезоксирибонуклеозид дифосфаты. Все ферменты редуктазы использоют общие сульфгидрильные радикального механизма зависит от реактивного цистеина остатков, которые окисляются с образованием дисульфидных связей в ход реакции. РНР фермент перерабатывается в результате реакции с тиоредоксином или глутаредоксином.
Регулирование РНР и связанных с ними ферментов, поддерживает баланс по отношению друг к другу. Очень низкая концентрация подавляет синтез ДНК и репарацию ДНК и является летальным для клетки, в то время как ненормальные отношения является мутагенным в связи с увеличением вероятности ДНК-полимеразы включения в процессе синтеза ДНК.
При синтезе нуклеиновых кислот РНК, аденозина, полученных от АТФ является одним из четырех нуклеотидов включены непосредственно в молекулах РНК, РНК-полимеразы. Энергия, это полимеризация происходит с отщеплением пирофосфата (две фосфатные группы). Этот процесс аналогичен в биосинтезе ДНК, за исключением того, что АТФ сводится к дезоксирибонуклеотиду дАТФ, перед включением в ДНК .
В синтезе белка . Аминоацил-тРНК-синтетазы используют ферменты АТФ в качестве источника нергии для прикрепления молекулы тРНК к его конкретной аминокислоте, образующей аминоацил-тРНК, готовый к переводом на рибосомы. Энергия становится доступной в результате гидролиза АТФ аденозинмонофосфата (AMФ) удаляют две фосфатные группы.
АТФ используется для многих клеточных функций, включая транспортную работу перемещения веществ через клеточные мембраны. Он также используется для механической работы, поставка энергии, необходимой для мышечного сокращения. Она поставляет энергию не только в сердечной мышце (для циркуляции крови) и скелетных мышц (например, за грубое движение тела), но также и к хромосомам и жгутиков, чтобы они могли выполнять свои многочисленные функции. Большую роль АТФ в химической работе, предоставлению необходимой энергии для синтеза нескольких тысяч типов макромолекул, что клетка должна существовать.
АТФ также используется в качестве включения-выключения как для контроля химических реакций и для отправки информации. Форма белковых цепей, которые производят строительные блоки и другие структуры, используемые в жизни определяется в основном слабые химические связи, которые легко исчезают и переструктурируются. Эти цепи могут сократить, удлинить, а также изменить форму в ответ на ввод или вывод энергии. Изменения в цепях изменить форму белка, а также может изменять свои функции или вызвать его, чтобы стать активным или неактивным.
Молекулы АТФ могут связываться в одной части молекулы белка, в результате чего другая часть той же молекулы, чтобы скользить или слегка двигаться который заставляет его изменять свою конформацию, инактивации молекул. После удаления АТФ вызывает белок, чтобы вернуться к своей первоначальной форме, и таким образом она снова функциональным.
Цикл может повторятся до тех пор, пока молекула возвращается, эффективно действующая в качестве и выключателя и переключателя. Оба добавлением фосфора (фосфорилирование) и удаления фосфора из белка (дефосфорилирование) может служить либо включения или выключения .
3.4 Другие функции АТФ
Роль в метаболизме, синтезе и активном транспорте
Таким образом, АТФ передает энергию между пространственно разделенных метаболических реакций. АТФ является основным источником энергии для большинства клеточных функций. Это включает в себя синтез макромолекул, включая ДНК и РНК, и белки. АТФ также играет важную роль в транспорте макромолекул через клеточные мембраны, например, экзоцитоза и эндоцитоза.
Роль в структуре клеток и передвижения
ATФ участвует в поддержании клеточной структуры путем облегчения монтажа и демонтажа элементов цитоскелета. В связи с этим процессом, АТФ, необходимых для сокращения нитей актина и миозина необходимых для мышечного сокращения. Этот последний процесс является одним из основных требований энергию животных и имеет важное значение для движения и дыхания.
Роль в сигнальных системах
Во внеклеточных сигнальных системах
АТФ также является сигнальной молекулой. АТФ, АДФ, или аденозин признаны пуринергическими рецепторов. Пуринорецепторы могут быть наиболее распространенных рецепторов в тканях млекопитающих.
У людей этой сигнализации роль важна как в центральной и периферической нервной системы. Активность зависит от выпуска АТФ из синапсов, аксонов и глии пуринергическими активирует рецепторы мембраны
Во внутриклеточных сигнальных системах
АТФ имеет решающее значение в передаче сигнала процессов. Он используется киназ в качестве источника фосфатных групп в их реакции фосфата передачи. Киназы на подложках, таких как белки или липиды мембраны являются распространенной формой сигнала. Фосфорилирование белка по киназе могут активировать этот каскад, такие как митогенактивированной протеинкиназыкаскада.
АТФ используется также аденилатциклазу и превращается в вторичный мессенджер молекулы АМФ, который участвует в запуске кальция сигналы высвобождение кальция из внутриклеточных депо. [ 38 ] Эта форма сигнала имеет особенно важное значение в функции мозга, хотя он участвует в регуляции множества других клеточных процессов .
Заключение
1. Аденозинтрифосфат - нуклеотид, играет исключительно важную роль в обмене энергии и веществ в организмах; в первую очередь соединение известно как универсальный источник энергии для всех биохимических процессов, протекающих в живых системах. Химически АТФ представляет собой трифосфорный эфир аденозина, который является производным аденина и рибозы. По строению АТФ сходна с адениновым нуклеотидом, входящим в состав РНК, только вместо одной фосфорной кислоты в состав АТФ входят три остатка фосфорной кислоты. Клетки не в состоянии содержать кислоты в заметных количествах, а только их соли. Поэтому фосфорная кислота входит в АТФ в виде остатка (вместо ОН-группы кислоты имеется отрицательно заряженный атом кислорода).
2. В организме АТФ синтезируется путём фосфорилирования АДФ:
АДФ + H 3 PO 4 + энергия > АТФ + H 2 O.
Фосфорилирование АДФ возможно двумя способами: субстратное фосфорилирование и окислительное фосфорилирование (используя энергию окисляющихся веществ).
Окислительное фосфорилирование - один из важнейших компонентов клеточного дыхания, приводящего к получению энергии в виде АТФ. Субстратами окислительного фосфорилирования служат продукты расщепления органических соединений - белки, жиры и углеводы. Процесс окислительного фосфорилирования проходит на кристах митохондрий.
Субстр атное фосфорил ирование (биохимическое), синтез богатых энергией фосфорных соединений за счёт энергии окислительно-восстановительных реакций гликолиза и при окислении a-кетоглутаровой кислоты в цикле трикарбоновых кислот.
3. Главная роль АТФ в организме связана с обеспечением энергией многочисленных биохимических реакций. Являясь носителем двух высокоэнергетических связей, АТФ служит непосредственным источником энергии для множества энергозатратных биохимических и физиологических процессов. В биоэнергетике живых организмов имеют значение: химическая энергия запасается путём образования АТФ, сопряжённого с экзергоническими катаболическими реакциями окисления органических субстратов; химическая энергия утилизируется путём расщепления АТФ, сопряжённого с эндергоническими реакциями анаболизма и другими процессами, требующими затраты энергии.
4. При усиленной нагрузке (например, в беге на короткие дистанции) мышцы работают исключительно за счет запаса АТФ. В клетках мышц этого запаса хватает на несколько десятков сокращений, а дальше количество АТФ должно восполняться. Синтез АТФ из АДФ и АМФ происходит за счет энергии, выделяющейся при расщеплении углеводов, липидов и других веществ. На выполнение умственной работы также затрачивается большое количество АТФ. По этой причине людям умственного труда требуется повышенное количество глюкозы, расщепление которой обеспечивает синтез АТФ.
Помимо энергетической АТФ выполняет в организме ещё ряд других не менее важных функций:
· Вместе с другими нуклеозидтрифосфатами АТФ является исходным продуктом при синтезе нуклеиновых кислот.
· Кроме того, АТФ отводится важное место в регуляции множества биохимических процессов. Являясь аллостерическим эффектором ряда ферментов, АТФ, присоединяясь к их регуляторным центрам, усиливает или подавляет их активность.
· АТФ является также непосредственным предшественником синтеза циклического аденозинмонофосфата - вторичного посредника передачи в клетку гормонального сигнала.
Также известна роль АТФ в качестве медиатора в синапсах.
Библиографический список
1. Лемеза, Н.А. Пособие по биологии для поступающих в ВУЗы / Л.В. Камлюк Н.Д. Лисов. - Мн.: Юнипресс, 2011 г. - 624 с.
2. Lodish, H, Berk A, Matsudaira P, Kaiser CA, Krieger M, Scott MP, Zipursky SL, Darnell J. Molecular Cell Biology, 5th ed. - New York: WH Freeman, 2004.
3. Романовский, Ю.М. Молекулярные преобразователи энергии живой клетки. Протонная АТФ-синтаза - вращающийся молекулярный мотор / Ю.М. Романовский А.Н. Тихонов // УФН. - 2010. - Т.180. - С.931 - 956.
4. Voet D, Voet JG. Biochemistry Vol 1 3rd ed. - Wiley: Hoboken, NJ. - N-Y: W. H. Freeman and Company, 2002. - 487 р.
5. Общая химия. Биофизическая химия. Химия биогенных элементов. М.: Высшая школа, 1993 г
6. Вершубский, А.В. Биофизика. / А.В. Вершубский, В.И. Прик-лонский, А.Н. Тихонов. - М: 471-481.
7. Альбертс Б. Молекулярная биология клетки в 3-х томах. / Альбертс Б., Брей Д., Льюис Дж. и др. М.: Мир, 1994.1558 с.
8. Николаев А.Я. Биологическая химия - М.: ООО "Медицинское информационное агенство", 1998.
9. Berg, J. M. Biochemistry, international edition. / Berg, J. M, Tymoczko, J. L, Stryer, L. - New York: WH Freeman, 2011; p 287.
10. Кнорре Д.Г. Биологическая химия: Учеб. для хим., биол. И мед. спец. вузов. - 3-е изд., испр. / Кнорре Д.Г., Мысина С.Д. - М.: Высш. шк., 2000. - 479 с.: ил.
11. Элиот, В. Биохимия и молекулярная биология / В. Элиот, Д. Элиот. - М.: Изд-во НИИ Биомедицинской химии РАМН, ООО "Материк-альфа", 1999, - 372 с.
12. Shina CL, K., 7 Areieh, W. On the Energetics of ATP Hydrolysis in Solution. Journal Of Physical Chemistry B,113 (47), (2009).
13. Berg, J. M. Biochemistry / J. M. Berg: J. L. Tymoczko, L. Stryer. - N-Y: W. H. Freeman and Company, 2002. - 1514 p.
...Подобные документы
Органические соединения в организме человека. Строение, функции и классификация белков. Нуклеиновые кислоты (полинуклеотиды), особенности строений и свойства РНК н ДНК. Углеводы в природе и организме человека. Липиды - жиры и жироподобные вещества.
реферат , добавлен 06.09.2009
Процесс синтеза белков и их роль в жизнедеятельности живых организмов. Функции и химические свойства аминокислот. Причины их нехватки в организме человека. Виды продуктов, в которых содержатся незаменимые кислоты. Аминокислоты, синтезируемые в печени.
презентация , добавлен 23.10.2014
Энергетическая, запасающая и опорно-строительная функции углеводов. Свойства моносахаридов как основного источника энергии в организме человека; глюкоза. Основные представители дисахаридов; сахароза. Полисахариды, образование крахмала, углеводный обмен.
доклад , добавлен 30.04.2010
Функции обмена веществ в организме: обеспечение органов и систем энергией, вырабатываемой при расщеплении пищевых веществ; превращение молекул пищевых продуктов в строительные блоки; образование нуклеиновых кислот, липидов, углеводов и других компонентов.
реферат , добавлен 20.01.2009
Роль и значение белков, жиров и углеводов для нормального протекания всех жизненно важных процессов. Состав, структура и ключевые свойства белков, жиров и углеводов, их важнейшие задачи и функции в организме. Основные источники данных пищевых веществ.
презентация , добавлен 11.04.2013
Характеристика структуры холестериновых молекул как важного компонента клеточной мембраны. Исследование механизмов регуляции обмена холестерина в организме человека. Анализ особенностей возникновения избытка липопротеидов низкой плотности в кровотоке.
реферат , добавлен 17.06.2012
Обмен белков, липидов и углеводов. Типы питания человека: всеядность, раздельное и низкоуглеводное питание, вегетарианство, сыроедение. Роль белков в обмене веществ. Недостаток жиров в организме. Изменения в организме в результате изменения типа питания.
курсовая работа , добавлен 02.02.2014
Рассмотрение участия железа в окислительных процессах и в синтезе коллагена. Ознакомление со значением гемоглобина в процессах кровообразования. Головокружения, одышка и нарушение обмена веществ как результат дефицита железа в человеческом организме.
презентация , добавлен 08.02.2012
Свойства фтора и железа. Суточная потребность организма. Функции фтора в организме, влияние, смертельная доза, взаимодействие с другими веществами. Железо в организме человека, его источники. Последствия дефицита железа для организма и его переизбытка.
презентация , добавлен 14.02.2017
Белки как источники питания, их основные функции. Аминокислоты, участвующие в создании белков. Строение полипептидной цепи. Превращения белков в организме. Полноценные и неполноценные белки. Структура белка, химические свойства, качественные реакции.





