Взаимодействие аллельных генов и его типы
Изучая законы Г.Менделя, мы уже знакомились с основными типами взаимодействия аллельных генов. На основе ранее изученного материала заполните таблицу.
Таблица. Типы взаимодействия генов одной аллельной пары
Тип взаимодействия |
Характер взаимодействия в |
Формулы расщепления по фенотипу в F 2 и при анализирующем скрещивании |
Примеры |
| Полное доминирование | Один аллель (доминантный) полностью подавляет действие другого (рецессивного) аллеля |
Высокий рост растения доминирует над карликовостью; вьющиеся волосы – над прямыми; карие глаза – над голубыми и т. п. |
|
| Неполное доминирование | Действие каждого аллеля реализуется не полностью, вследствие чего проявляется промежуточный признак |
Окраска цветков ночной красавицы; серповидноклеточная анемия; цистонурия и т.п. |
|
| Кодоминирование | Участие обоих аллелей в определении признака |
IV группа крови (эритроциты этой группы крови имеют агглютиногены типа А и типа В ) |
Взаимодействие неаллельных генов
Исходя из законов Г.Менделя, можно прийти к заключению, что существует довольно прочная связь между геном и признаком, что генотип слагается из суммы независимо действующих генов, а фенотип – механическая совокупность отдельных признаков. Однако прямые и однозначные связи гена с признаком скорее исключение, чем правило. Развитие признака организма обычно находится под контролем многих генов, и признак является результатом взаимодействия неаллельных генов.
Простейший случай взаимодействия неаллельных генов – когда признак контролируется двумя парами аллелей. Рассмотрим некоторые примеры такого взаимодействия.
Комплементарное взаимодействие генов
Явление взаимодействия генов, при котором для проявления признака необходимо наличие двух неаллельных генов, называют комплементарностью (от лат. complementum – дополнение), а гены, необходимые для проявления признака, комплементарными , или дополнительными.
Рассмотрим различные варианты комплементарного взаимодействия генов.
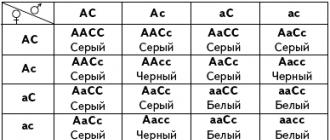
Аллели второго гена проявляются только при наличии доминантного аллеля гена А . Окраска мышей зависит в простейшем случае от двух генов. Мыши с генотипом аа лишены пигмента и имеют белый цвет. При наличии доминантного аллеля А пигмент вырабатывается, и мышь как-то окрашена. Конкретный цвет определяется вторым геном. Его доминантный аллель С обусловливает серый цвет мыши, а рецессивный аллель с – черный цвет. Таким образом, если рассматривать гомозиготные варианты, генотип серых мышей – ААСС , черных – ААсс , белых – ааСС или аасс . При скрещивании серой мыши ААСС с белой аасс в первом поколении все получается по Менделю: все гибриды имеют серый цвет (это гетерозиготы с генотипом АаСс ). В F2, как легко проверить, получим 9/16 серых мышей, 3/16 черных и 4/16 белых.

Для проявления признака в генотипе должны присутствовать доминантные аллели двух разных генов. Примером такого комплементарного взаимодействия генов является наследование формы плода у тыквы. При наличии обоих доминантных аллелей плоды имеют дисковидную форму, при наличии одного (любого!) доминантного аллеля – сферическую, а при отсутствии доминантных аллелей – удлиненную.

Для проявления признака в генотипе должны присутствовать доминантные аллели двух разных генов, но каждый доминантный аллель в сочетании с рецессивными аллелями другой пары имеет самостоятельное фенотипическое проявление. Например, у кур гороховидная форма гребня определяется одним доминантным геном, розовидная – другим неаллельным ему, но тоже доминантным геном. Когда эти гены окажутся в одном генотипе, развивается ореховидная форма гребня. В случае если организм оказывается гомозиготным по обоим рецессивным генам, развивается простой листовидный гребень. При скрещивании дигибридов (все с ореховидным гребнем) во втором поколении происходит расщепление в отношении 9:3:3:1. Но здесь нельзя найти независимого расщепления каждого аллеля в отношении 3:1, так как во всех случаях совпадения в генотипе обоих доминантных генов их прямого действия не обнаруживается.
Таким образом, о комплементарном взаимодействии неаллельных генов говорят в том случае, когда два неаллельных гена дают новый признак, то есть имеет место новообразование.
Эпистатическое взаимодействие генов
Взаимодействие генов, при котором один ген подавляет действие другого, неаллельного первому, называется эпистазом (от греч. эпи – над и стазис – стоять), а гены, подавляющие действие других генов, называются генами-супрессорами , или генами-ингибиторами (от лат. inhiber – удерживать), или эпистатическими генами . Подавляемый ген называется гипостатическим .
Различают эпистаз доминантный и рецессивный.
Доминантный эпистаз связан с доминантным геном-ингибитором. Например, ген I обусловливает белый цвет плодов тыквы, при его наличии действие гена В не проявляется. При генотипе iiВВ или iiВb плоды тыквы имеют желтый цвет. Наконец, если оба гена представлены рецессивными аллелями, плоды тыквы имеют зеленый цвет. По этому же типу может наследоваться окраска шерсти собак, овец и других животных.
Рецессивный эпистаз обнаруживается тогда, когда проявление гена подавляется рецессивными аллелями другого гена. Этот вид эпистаза иллюстрируется наследованием окраски шерсти у домовых мышей. Окраска агути (рыжевато-серая) определяется доминантным геном А , его рецессивный аллель а дает черную окраску. Ген из неаллельной пары В способствует проявлению цветности, а ген b является супрессором и подавляет действие доминантного аллеля А и рецессивного а . Мыши с генотипом Аbb неотличимы по фенотипу от особей с генотипом ааbb – все белые.
Полимерное взаимодействие генов
Полимерия – тип взаимодействия неаллельных генов, при котором несколько пар неаллельных генов действуют на развитие одного признака. Такие гены называются полимерными (от греч. полис – много и мерос – часть). Их обозначают одной буквой, но с разными индексами, которые указывают на число аллельных пар в генотипе, обусловливающих развитие конкретного признака (А 1 А 1 А 2 А 2 ).
Различают два варианта полимерии: с суммирующим действием генов и без усиления генов друг другом.
Суммирующее действие полимерных генов. В этом случае степень проявления признака зависит от числа доминантных аллелей в генотипе особи. Так, красная окраска зерен пшеницы определяется двумя и более парами генов. Каждый из доминантных генов этих аллелей определяет красную окраску, рецессивные гены определяют белый цвет зерен. Один доминантный аллель дает не очень сильно окрашенные зерна. Если в генотипе присутствуют два доминантных аллеля, интенсивность окраски повышается. Лишь в том случае, когда организм оказывается гомозиготным по всем парам рецессивных генов, зерна не окрашены. Таким образом, при скрещивании дигибридов происходит расщепление в отношении 15 окрашенных к одному белому. Но из 15 окрашенных один будет иметь интенсивный красный цвет, т.к. содержит четыре доминантных аллеля, четыре будут окрашены несколько светлее, т.к. содержат в генотипах три доминантных аллеля и один рецессивный аллель, шесть – еще светлее с двумя доминантными и двумя рецессивными аллелями в генотипах, четыре – еще более светлые, т.к. имеют лишь один доминантный и три рецессивных гена, то есть истинное расщепление будет 1:4:6:4:1.
По данному варианту полимерии наследуются окраска кожи, рост и масса у человека. Подобный же механизм наследования характерен для многих количественных, в том числе и хозяйственно-ценных признаков: содержание сахара в корнеплодах свеклы, содержание витаминов в плодах и овощах, длина колоса злаков, длина початка кукурузы, плодовитость животных, молочность скота, яйценоскость кур и др.
Полимерные гены не усиливают друг друга. В этом случае расщепление в F 2 будет 15:1. Так, плоды пастушьей сумки могут быть треугольными (доминантный признак) и овальными (рецессив). Признак контролируется двумя парами полимерных генов. Если в генотипе растения имеется хотя бы один доминантный аллель из первой или второй пары полимерных генов, то форма плода у него будет треугольной (А 1 А 2 ; А 1 а 2 ; а 1 А 2 ). Овальную форму плодов будут иметь лишь те растения, у которых в генотипе нет ни одного доминантного аллеля – а 1 а 1 а 2 а 2 .
Таким образом, накопление определенных аллелей в генотипе может привести к изменению выраженности признаков.
Множественное действие генов
 Часто ген оказывает действие не на один, а на ряд признаков организма. Явление, при котором один ген может влиять на формирование нескольких признаков организма, называется плейотропией
(от греч. плеон
– более многочисленный и тропос
– поворот).
Часто ген оказывает действие не на один, а на ряд признаков организма. Явление, при котором один ген может влиять на формирование нескольких признаков организма, называется плейотропией
(от греч. плеон
– более многочисленный и тропос
– поворот).
Существование этого явления отнюдь не противоречит классической концепции «один ген – один белок – один признак». Упрощенно влияние одного гена сразу на несколько признаков можно представить следующим образом. В результате считывания информации с гена образуется некий белок, который затем может участвовать в различных процессах, происходящих в организме, оказывая таким образом множественное действие. Например, у гороха бурая окраска кожуры семян и развитие пигмента в других частях растения, окраска цветков зависят от одного гена; у дрозофилы ген, обусловливающий белую окраску глаз, влияет на осветление окраски тела и изменение некоторых внутренних органов; у человека ген, отвечающий за рыжий цвет волос, одновременно определяет более светлую окраску кожи и появление веснушек. Синдром Марфана обусловлен аутосомным доминантным плейотропным геном и проявляется высоким ростом, удлинением костей пальцев рук и ног (паучьи пальцы), гиперподвижностью суставов, подвывихом хрусталика глаза, пороком сердца.
Такое множественное действие характерно для большинства генов. Однако не следует представлять, что плейотропный ген в равной степени влияет на каждый из признаков. Для абсолютного большинства генов с той или иной степенью плейотропии характерно более сильное влияние на один признак и значительно более слабое – на другой.
Генотип как целостная система
Факт расщепления в потомстве гибридов позволяет утверждать, что генотип слагается из отдельных элементов – генов, которые могут наследоваться независимо (дискретность генотипа ). В то же время генотип не может рассматриваться как простая механическая сумма отдельных генов. Генотип – это система взаимодействующих генов. Точнее, взаимодействуют не сами гены (участки ДНК), а образуемые на их основе генные продукты (РНК, а затем – белки). Поэтому в отдельных случаях действие разных генов относительно независимо, но, как правило, проявление признака есть результат взаимодействия продуктов разных генов.
Генотип любого организма представляет собой сложную целостную систему взаимодействующих генов. Эта целостность генотипа возникла исторически в процессе эволюции вида. В результате мутаций постоянно появляются новые гены, формируются новые хромосомы и даже новые геномы. Вновь возникшие гены могут сразу же взаимодействовать с уже имеющимися генами или модифицировать характер работы последних, даже будучи рецессивными, то есть, не проявляясь сами по себе.
Следовательно, у каждого вида растений и животных генотип проявляет себя как исторически сложившаяся к данному моменту целостная система.
II. Закрепление знаний
Обобщающая беседа по ходу изучения нового материала.
III. Домашнее задание
Изучить параграф учебника (связь между генами и признаками, типы взаимодействия аллельных и неаллельных генов, плейотропия, генотип как целостная система).
Решить задачи в конце параграфа учебника.
Другим типом взаимодействия неаллельных генов является комплементарность. Она заключается в том, что развитие признака требует наличия в генотипе доминантных аллелей двух определенных генов. Классическим примером комплементарного взаимодействия генов является наследование окраски лепестков венчика цветов душистого горошка. При скрещивании цветов белой окраски у потомства появляется новый признак – лепестки венчика красной окраски, а во втором поколении расщепление составляет 9 красных к 7 белых.
М – хромоген N – хромогеназа
m – отсутствие n – отсутствие
Р: ♀ ММnn ´ ♂ mmNN
белые белые
по генотипу: дигетерозиготны
по фенотипу: пурпурно – красные
P: ♀ MnNn ´ ♂ MmNn
F 2: по решётке Пеннета
| ♀ ♂ | MN | Mn | mN | mn |
| MN | MMNN | MMNn | MmNN | MmNn |
| Mn | MMNn | MMnn | MmNn | Mmnn |
| mN | MmNN | MmNn | mmNN | mmNn |
| mn | MmNn | Mmnn | mmNn | mmnn |
по генотипу: 1: 2: 2: 1: 4: 1: 2: 2: 1
по фенотипу: 9: 7
пурпурно - красные белые
Таким образом, при комплементарном взаимодействии генов также наблюдается отклонение от закона независимого наследования.
У человека комплементарным действием обладают гены пигментации волос:
m 1 – значительное количество меланина
m 2 - среднее количество меланина
m 3 – малое количество меланина
R - красный пигмент
r - отсутствие пигмента
Сочетание аллелей указанных генов дают весь спектр окрасок волос. При этом степень доминирования следующая: тm 1 >m 2 >R>m 1 >r
Генотипы: Фенотип:
m 1 m 1 RR брюнет (с глянцем)
m 1 m 1 Rr брюнет (лоснящиеся волосы)
m 1 m 1 rr брюнет
m 1 m 2 RR темный шатен
m 1 m 3 rr шатен
m 2 m 2 Rr каштановый
m 2 m 2 RR темно-рыжий
М 2 m 3 RR темно-рыжий
m 3 m 3 RR ярко-рыжий
m 3 m 3 Rr блондин с рыжеватым оттенком
m 3 m 3 rr блондин
Другим примером комплементарного взаимодействия является продукция клетками человека противовирусного вещества – интерферона. Его синтез зависит от присутствия в генотипе двух доминантных генов из разных аллельных пар:
Фенотипический радикал: Фенотип:
А-В - интерферон синтезируется
ааВ – интерферон не синтезируется
А-вв интерферон не синтезируется
аавв интерферон не синтезируется
Наследование нормального гемоглобина зависит от 4-х доминантных генов из разных аллельных пар. Только при фенотипическом радикале А-В-С-Д- гемоглобин связывается с О 2 (оксигемоглобин) и с СО 2 (карбоксигемоглобин). При всех других сочетаниях генов как-то.
Комплементарным называется такой вид взаимодействия неаллельных генов, при котором действие гена из одной аллельной пары дополняется действием гена из другой аллельной пары, в результате чего формируется качественно новый признак.
Классический пример такого взаимодействия – наследование формы гребня у кур. Встречаются следующие формы гребня: листовидный – результат взаимодействия двух рецессивных неаллельных генов а abb ; ореховидный – результат взаимодействия двух доминантных неаллельных генов A - B -; розовидный и гороховидный – c генотипами A - bb и aaB - , соответственно.
Другой пример – наследование окраски шерсти у мышей. Окраска бывает серая, белая и черная, а пигмент только один – черный. В основе формирования той или иной окраски шерсти лежит взаимодействие двух пар неаллельных генов:
A – ген, определяющий синтез пигмента;
a – ген, не определяющий синтез пигмента;
B – ген, определяющий неравномерное распределение пигмента;
b – ген, определяющий равномерное распределение пигмента.
Примеры комплементарного взаимодействия у человека: ретинобластома и нефробластома кодируются двумя парами неаллельных генов.
Возможные варианты расщепления в F 2 при комплементарном взаимодействии: 9:3:4; 9:3:3:1; 9:7.
Эпистаз
Эпистаз - такой вид взаимодействия неаллельных генов, при котором действие гена из одной аллельной пары подавляется действием гена из другой аллельной пары.
Различают две формы эпистаза – доминантный и рецессивный. При доминантном эпистазе в качестве гена-подавителя (супрессора) выступает доминантный ген, при рецессивном эпистазе – рецессивный ген.
Пример доминантного эпистаза – наследование окраски оперения у кур. Взаимодействуют две пары неаллельных генов:
С – ген, определяющий окраску оперения (обычно пеструю),
с – ген, не определяющий окраску оперения,
I – ген, подавляющий окраску,
i – ген, не подавляющий окраску.
Варианты расщепления в F 2: 12:3:1, 13:3.
У человека примером доминантного эпистаза являются ферментопатии (энзимопатии) – заболевания, в основе которых лежит недостаточная выработка того или иного фермента.
Пример рецессивного эпистаза – так называемый «бомбейский феномен»: в семье у родителей, где мать имела группу крови О, а отец – группу крови А, родились две дочери, из которых одна имела группу крови АВ. Ученые предположили, что у матери в генотипе был ген I B , однако его действие было подавлено двумя рецессивными эпистатическими генами dd.
Полимерия
Полимерия - такой вид взаимодействия неаллельных генов, при котором несколько неаллельных генов определяют один и тот же признак, усиливая его проявление. Это явление противоположно плейотропии. По типу полимерии обычно наследуются количественные признаки, чем и обусловлено большое разнообразие их проявления в природе.
Например, окраска зерен у пшеницы определяется двумя парами неаллельных генов:
A 1
a 1 – ген, не определяющий красную окраску;
A 2 – ген, определяющий красную окраску;
a 2 – ген, не определяющий красную окраску.
A 1 A 1 A 2 A 2 – генотип растений с красной окраской зерен;
a 1 a 1 a 2 a 2 - генотип растений с белой окраской зерен.
Расщепление в F 2: 15:1 или 1:4:6:4:1.
У человека по типу полимерии наследуются такие признаки, как рост, цвет волос, цвет кожи, величина артериального давления, умственные способности.
Основные формы взаимодействия неалельных генов – комплементарность, эпистаз и полимерия. Они преимущественно видоизменяют классическую формулу расщепления за фенотипом, установленную Г. Менделем для дигибридного скрещивания (9:3:3:1).
Комплементарность (лат. complementum – дополнения). Комплементарными, или взаимодополняющими, называются неаллельные гены, которые поодиночке не проявляют своего действия, но при одновременном наличии в генотипе предопределяют развитие нового признака. У душистого горошка окраски цветков обусловлена двумя доминантными неаллельными генами, из них один ген (А) обеспечивает синтез бесцветного субстрата, другой (В) – синтез пигмента. Поэтому при скрещивании растений с белыми цветками (ААbb х ааВВ) все растения в первом поколении F1 (АаВb) имеют окрашенные цветки, а во втором поколении F2 происходит расщепления за фенотипом в соотношении 9:7, где 9/16 растений имеют окрашенные цветки и 7/16 – неокрашенные.
У человека нормальный слух обусловлен комплементарным взаимодействием двух доминантных неаллельных генов D и Е, из них один определяет развитие завитка, другой – слухового нерва. Люди с генотипами D–Е– имеют нормальный слух, с генотипами D–ее и ddЕ– – глухие. В браке, где родители глухие (DDee ´ ddEE), все дети будут иметь нормальный слух (DdEe).
Епистаз – взаимодействие неаллельных генов, при котором один ген подавляет действие другого, неаллельного, гена. Первый ген называется эпистатическим, или супрессором (ингибитором), другой, неаллельний, ген – гипостатическим. Если эпистатический ген – доминантный, эпистаз называют доминантным (А>В). И, наоборот, если эпистатический ген рецессивный, эпистаз – рецессивный (аа>В или аа >вв). Взаимодействие генов при эпистазе противоположно комплементарности.
Пример доминантного эпистаза . У кур доминантный аллель С одного гена обуславливает развитие окраски перья, но доминантный аллель І другого гена является его супрессором. Поэтому куры с генотипом І–С– – белые, а с генотипами ііСС и ііСс – окрашенные. В скрещивании белых кур (ІІСС х іісс) гибриды первого поколения F1 окажутся белыми, но при скрещивании F1 между собой во втором поколении F2 состоится расщепления за фенотипом в соотношении 13:3. Из 16 особей 3 будут окрашены (ііСС и ііСс), так как в них отсутствует доминантный ген-супрессор и есть доминантный ген окраски. Другие 13 особей будут белыми.
Примером рецесивного эпистаза может быть бомбейский феномен – необыкновенное наследование групп крови системы АВО, впервые выявленное в одной индийской семье. В семье, где отец имел группу крови І (О), а иметь – ІІІ (В), родилась девочка с группой І (О), она вступила в брак с мужчиной с группой крови ІІ(А) и у них родилось две девочки: одна из группой крови ІV (АВ), другая – с І (О). Рождение девочки с ІV (АВ) группой крови в семье, где отец имел ІІ (А), а мама – І (О) было необыкновенным. Генетики объяснили этот феномен так: девочка с группой ІV (АВ) унаследовала аллель ІА от отца, а аллель ІВ – от матери, но у матери аллель ІВ фенотипически не проявлялся, так как в ее генотипе присутствовал редкий рецессивний эпистатический ген s в гомозиготном состоянии, который спровоцировал фенотипичное проявление аллеля ІВ.
Гипостаз – взаимодействие неалельных генов, при котором доминантный ген одной аллельной пары подавляется эпистатическим геном из другой аллельной пары. Если ген А подавляет ген В (А>В), то по отношению к гену В взаимодействие неаллельных генов называется гипостазом, а по отношению к гену А – эпистазом.
Полимерия – взаимодействие неаллельных генов, при котором один и и тот же признак контролируют несколько доминантных неаллельных генов, которые действуют на этот признак однозначно, в равной степени, усиливая его проявление. Такие однозначные гены называют полимерными (множественными, полигенами) и их обозначают одной буквой латинского алфавита, но с разными цифровыми индексами. Например, доминантные полимерные гены – А1, А2, А3 и т.д., рецессивные – а1, а2, а3 и т.д. Соответственно обозначают генотипы – А1А1А2А2А3А3, а1а1а2а2а3а3. Признаки, которые контролируются полигенами, называют полигенными, а наследования этих признаков – полигенным, в отличие от моногенного, где признак контролируется одним геном. Явление полимерии впервые описал в 1908 г. шведский генетик Г. Нильсон-Эле при изучении наследования цвета зерна пшеницы.
Полимерия бывает кумулятивной и некумулятивной . При кумулятивной полимерии каждый ген в отдельности имеет слабое действие (слабую дозу), но количество доз всех генов в конечном результате суммируется, так что степень выражения признака зависит от числа доминантных аллелей. За типом полимерии у человека наследуются рост, масса тела, цвет кожи, умственные способности, величина артериального давления. Так, пигментация кожи у человека определяется 4-6 парами полимерных генов. В генотипе коренных жителей Африки имеются преимущественно доминантные аллели (Р1Р1Р2Р2Р3Р3Р4Р4), у представителей европеоидной рассы – рецессивные (p1p1p2p2p3p3p4p4). От брака темнокожего и белой женщины рождаются дети с промежуточным цветом кожи – мулаты (Р1р1P2р2P3р3P4р4). Если супруги – мулаты, то возможное рождение детей с пигментацией кожи от максимально светлой к максимально темной.
Полигенно в типичных случаях наследуются количественные признаки. Тем не менее в природе существуют примеры полигенного наследования качественных признаков, когда конечный результат не зависит от числа доминантных аллелей в генотипе – признак или проявляется, или не проявляется (некумулятивная полимерия).
Плейотропия – способность одного гена контролировать несколько признаков (множественное действие гена). Так, синдром Марфана в типичных случаях характеризуется триадой признаков: подвывихом хрусталика глаза, пороками сердца, удлинением костей пальцев рук и ног (арахнодактилия – паучьи пальцы). Этот комплекс признаков контролируется одним аутосомно-доминантным геном, который вызывает нарушения развития соединительной ткани.
Комплементарность – тип взаимодействия двух доминантных неаллельных генов, при котором один из них дополняет действие другого, и они вместе определяют новый признак, который отсутствовал у родительских особей, причём данный признак развивается только в присутствии обоих генов. Примером может служить серый окрас шерсти у мышей, который контролируется двумя генами (А и В), первый из которых детерминирует синтез пигмента (особи с генотипами АА и Аа – альбиносы), а второй - его накопление у основы и на кончиках волос. При скрещивании дигетерозигот получаем гибриды в соотношении 9:3:4 (серые:черные:белые), т.е., серыми будут особи, в генотипе которых будет присутствовать сочетание аллелей АВ, черными – особи с сочетанием аллелей аа, белыми – bb. У человека по данному принципу происходит синтез белка интерферона.
Эпистаз – взаимодействие неаллельных генов, при котором действие одного из них угнетает действие другого. В зависимости от того, какой из генов вызывает угнетение, выделяют доминантный и рецессивный эпистаз. Угнетающий ген получил название ингибитора, супрессора или же эпистатического гена, угнетаемый – гипостатического гена. Гены-ингибиторы в основном угнетают другой ген, не детерминируя развитие определенного признака. Расщепление по фенотипу составляет 13:3, 12:3:1, 9:3:4 и т.д. Если ген-супрессор рецессивный, то может наблюдаться криптомерия – зависимость признака от нескольких действующих одновременно генов, каждый из которых не имеет видимого фенотипического проявления.
Полимерное наследование признаков у человека. Плейотропия.
Одинаковое действие двух и более неаллельных генов на развитие одного и того же признака в организме называется полимерным взаимодействием генов. Полимерное взаимодействие проявляется в количественных признаках: рост, вес, окраска кожных покровов, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови, особенности нервной системы, уровень интеллекта. Степень развития количественных признаков зависит от числа воздействующих полимерных генов.
Первоначально полимерию было путем скрещивания сортов пшеницы с красными (A1A1A2A2) и белыми (а1а1а2а2) зернами. Получились растения F1, зерна которых были розового цвета. Скрестив гибриды F1, он получил растения F2, которые можно разделить на пять групп по цвету их зерен. Количественное соотношение их было следующим: растения с красным зерном - 1, с бледно-красным зерном - 4, с розовым зерном - 6, с бледно-розовым зерном - 4, с белым зерном - 1.
Примером полимерного наследования у человека является наследование окраски кожных покровов. В браке индивида негроидной расы с черной окраской кожи и представителем европеоидной расы с белой кожей дети рождаются с промежуточным цветом кожи (мулаты). В браке двух мулатов потомки могут обладать любой окраской кожи: от черной до белой, поскольку пигментация кожи обусловлена действием трех или четырех неаллельных генов. Влияние каждого из этих генов на окраску кожи примерно одинаково.
Плейотропия - явление множественного действия гена. Выражается в способности одного гена влиять на несколько фенотипических признаков. Таким образом, новая мутация в гене может оказать влияние на некоторые или все связанные с этим геном признаки. Этот эффект может вызвать проблемы при селективном отборе, когда при отборе по одному из признаков лидирует один из аллелей гена, а при отборе по другим признакам - другой аллель этого же гена.
Виды плейотропии
1) Первичная : ген одновременно проявляет множественное действие. Например, синдром Марфана обусловлен действием одного гена. Этот синдром проявляется следующими признаками: высокий рост за счет длинных конечностей, тонкие пальцы, порок сердца, высокий уровень катехоламинов в крови. Другим примером у человека служит серповидноклеточная анемия. Мутация нормального аллеля ведёт к изменению молекулярной структуры белка гемоглобина, при этом эритроциты теряют способность транспорту кислорода и приобретают серповидную форму вместо округлой. Гомозиготы по гену серповидноклеточности гибнут при рождении, гетерозиготы живут и обладают устойчивостью против малярийного плазмодия. Доминантная мутация, вызывающая у человека укорочение пальцев (брахидактилия), в гомозиготном состоянии приводит к гибели эмбриона на ранних стадиях развития.
2) Вторичная : имеется одно первичное фенотипическое проявление гена, которое обуславливает проявление вторичных признаков. Например, аномальный гемоглобин S в гомозиготном состоянии фенотипически первично проявляется в виде серповидноклеточной анемии, которая приводит к вторичным фенотипическим проявлениям в виде невосприимчивости к малярии, анемии, поражению сердца и мозга.
Примеры:
● Ген рыжих волос обусловливает более светлую окраску кожи и появление веснушек.
● Фенилкетонурия (ФКУ), болезнь, вызывающая задержку умственного развития, выпадение волос и пигментацию кожи, может быть вызвана мутацией в гене, кодирующем фермент фенилаланин-4-гидроксилаза, который в норме катализирует превращение аминокислоты фенилаланина в тирозин.
● Рецессивная мутация в гене, кодирующем синтез глобиновой части в гемоглобине (замена одной аминокислоты), вызывающая серповидную форму эритроцитов, изменения в сердечно-сосудистой, нервной, пищеварительной и выделительной системах.
● Арахнодактилия, вызываемая доминантной мутацией, проявляется одновременно в изменениях пальцев рук и ног, вывихах хрусталика глаза и врождённых пороках сердца.
● Галактоземия, вызываемая рецессивной мутацией гена, кодирующего фермент галактозо-1-фосфатуридилтрансфераза, приводит к слабоумию, циррозу печени и слепоте.
43. Сцепленное наследования генов (закон Т. Моргана). Кроссинговер. Генетические и цитологические карты хромосом.
Сцепленное наследование генов (Закон Т.Моргана): гены, расположенные в одной хромосоме, наследуются совместно - сцепленно, то есть наследуются преимущественно вместе. Полное сцепление - разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным. Неполное сцепление - разновидность сцепленного наследования, при которой гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Кроссинговер (от англ. Crossing-over - пере-крест) - обмен участками гомологичных хромосом в процессе клеточного деления, преимущественно в профазе первого мейотического деления, иногда в митозе.
Некроссоверные гаметы - гаметы, в процессе образования которых кроссинговер не произошел.
Нерекомбинанты - гибридные особи, у которых такое же сочетание признаков, как и у родителей.
Рекомбинанты - гибридные особи, имеющие иное сочетание признаков, чем у родителей.
Расстояние между генами измеряется в морганидах - условных единицах, соответствующих проценту кроссоверных гамет или проценту рекомбинантов. Например, расстояние между генами серой окраски тела и длинных крыльев (также черной окраски тела и зачаточных крыльев) у дрозофилы равно 17%, или 17 морганидам.
Генетическая карта - это отрезок прямой, на которой обозначены порядок расположения генов и указано расстояние между ними в процентах кроссинговера. Она строится на основе результатов анализирующего скрещивания. Картирование проводится, чтобы узнать, в какой паре хромосом и на какой генетической расстояния (коэффициент рекомбинации), или в какой части хромосомы расположены гены.
Цитологические карты хромосом - схематическое изображение хромосом с указанием мест фактического размещения отдельных генов, полученное с помощью цитологических методов. Цитологические карты хромосом составляют для организмов, для которых обычно уже имеются генетические карты хромосом. Каждое место расположения гена (локус) на генетической карте организма, установленное на основе частоты перекреста участков хромосом (кроссинговера) , на Цитологические карты хромосом привязано к определённому, реально существующему участку хромосомы, что служит одним из основных доказательств хромосомной теории наследственности.